緑のカエル。 カエルの内部構造 カエルの心臓の構造とその働き
両生類は、水と陸上の両方で繁栄する冷血の捕食動物です。 最初はえら呼吸をしますが、成体になると肺呼吸に切り替わります。 この記事では、カエルを例にして両生類の内部構造を詳しく調べます。
生息地
両生類は陸上と水中の 2 つの環境で生活し、よくジャンプしてよく泳ぎ、さらには木にも登ります。 その特性のおかげで、彼らは湿った場所(沼地、湿った森林、牧草地)と淡水域の岸辺の両方で繁栄します。 開発プロセス全体は水中で行われます。 そこで彼らは繁殖し、幼虫の発育と稚魚の成長が起こり、陸上では成熟した個体だけが見られます。
カエルの行動は環境の湿度にも依存します。 彼らは晴れた天候を許容せず、夕方や雨の日には狩りに行きます。 水の中やその近くに住んでいる動物は、日中も食べ物を探します。 寒さが始まると、動物は貯水池の底のシルトに潜り、寒い季節全体をそこで過ごします。 彼らは皮膚を通して呼吸できるので、水面に上がる必要はありません。 一部の動物は冬の間、落ち葉や大きな石の山の下に穴を掘って地表で過ごします。 体内のすべてのプロセスが遅くなり、暖かさが到着して初めて、たとえ凍った状態からでも通常の生活に戻ります。
カエルの外部構造の特徴
学童は通常、7 年生でカエルの内部構造を勉強します。 ただし、最初に外部構造を理解しましょう。 カエルの体は長さ8mm~32cmの頭と胴体から構成され、色は単色(緑、茶色、黄色)または斑入りの場合があります。 頸部領域は顕著ではなく、頭はすぐに体に入ります。 この動物は前肢と後肢が発達しています。 皮膚はむき出しで粘膜があり、角質はあまり発達していません。 表皮には、皮膚を乾燥から守る粘液物質を生成する多細胞腺が多数含まれています。 通常、5 本指タイプの地上の四肢は複雑な筋肉構造を持っています。 後肢は、特殊な運動方法により、肩、前腕、手からなる前肢よりも発達しています。 足の指は 4 本あり、男性の場合は内側の指の付け根に腫れがあり、これが生殖器疣贅です。 長い後肢は大腿部、下肢、足で構成され、遊泳膜で接続された 5 本の指があります。
カエルの頭
フラットヘッドには次のものがあります。

カエルの外部構造と内部構造
すべての両生類と同様に、カエルは長期間水がなくても大丈夫ですが、繁殖するには水が必要です。 変化した幼虫は魚らしさを失い、両生類に変わります。 体は細長く、四肢は二対あります。 魚と違って、体の中に入る頭は回転することができます。 骨格は骨で構成されていますが、軟骨もたくさんあります。 背骨には多くの椎骨があります。 肋骨がないということは、胸がないということです。 強い骨格と発達した筋肉のおかげで、この動物は陸上での生活に適応しています。 後肢と前肢には 3 つの関節があります。 皮膚は滑らかで、潤いを与える腺がたくさんあります。 カエルは肺と皮膚で呼吸します。

カエルの内臓の構造は、1 つの心室と 2 つの心房、および 2 つの循環円からなる 3 室の心臓の存在を示唆しています。 食物は咽頭から食道、胃、短腸を通過します。 それを消化するために、肝臓、胃壁、膵臓から分泌物が生成されます。 直腸の端には総排泄腔があり、そこに女性の卵管が開口しています。 動物には腎臓が 2 つと膀胱が 1 つあります。 小さな脳箱には、発達した前脳と小脳が含まれています。 カエルには視覚、聴覚、触覚、味覚、嗅覚の器官があります。
カエルの内部構造
筋肉はかなり複雑な構造をしており、魚に比べて非常によく発達しています。 筋肉群の協調的な働きのおかげでカエルは動くことができ、さらに呼吸にも関与します。

スケルトンには、脊椎、ガードル、四肢のスケルトン、頭蓋骨のセクションが含まれます。 後者は頸椎を使用して脊椎に接続されています。 これにより、首を傾けることが可能になります。 胴体には椎骨が7つあり、肋骨はありません。 仙骨領域は、頸部領域と同様に、1 つの椎骨で表されます。 長骨は尾の部分を形成します。 太もも、脚、足は後肢を形成し、肩、前腕、手は前肢を形成します。 それらは、前部と後部の四肢の帯を使用して脊椎に接続されています。 1 つ目は 2 つの肩甲骨と胸骨を含み、2 つ目は互いに融合した骨盤骨を含みます。
神経系
カエルの神経系は魚の神経系よりも複雑です。 その内部構造は次のとおりです: 神経、脊髄、脳。 後者は 3 つのセクションに分かれています。魚に比べてより発達した前脳、カエルは座ってばかりの生活を送り、単調な動きしかしないため小さな小脳、そして大脳半球です。 成人は上まぶたと下まぶた、および瞬膜を発達させており、そのおかげで角膜は乾燥せず、汚染から保護されています。
循環系
循環系は 3 室の心臓で表されます。 肺から動脈血が左心房に入ります。 右心房は内臓からの静脈血と真皮からの動脈血を受け取ります。

心房の収縮と同時に血液が心室に入ります。 特別な弁の助けを借りて、静脈血は肺と皮膚に流れ、動脈血は脳と頭部臓器に流れます。 混合血液は体の一部だけでなく、他のすべての臓器にも流れます。 カエルには 2 つの血液循環があり、それらは共通の心室によって結合されています。
呼吸器系
皮膚は呼吸に関与し、カエルの内部構造は血管網を持つ肺の助けを借りて呼吸を可能にします。

カエルは鼻孔を開き、口腔咽頭腔の底が下がり、空気が口腔に入ります。 次に、鼻孔が閉じて底が上がり、空気が肺に入ります。 肺の壁が崩壊し、腹筋が収縮すると、呼気が発生します。
消化器系
それはかなり大きな口腔咽頭腔から始まります。 カエルは獲物を見つけると舌を出し、獲物が舌にくっつきます。 上顎には小さな歯があり、獲物を捕まえる役割を果たします。 カエルの内臓の構造と活動は、食物の処理に貢献しています。 口腔咽頭腔内の唾液腺の分泌物によって湿らせられ、食道に入り、その後胃に入ります。 不完全に消化された食物は十二指腸に入り、次に小腸に入り、そこで栄養素が吸収されます。 未消化の残留物は、最初に直腸(後腸)を通過した後、総排出腔を通って出ます。
排泄系
仙骨の側面には糸球体を含み、血液から老廃物や一部の栄養素をろ過する腎臓が 2 つあります。

後者は尿細管に吸収されます。 尿は、まず尿管と総排出腔を通過して膀胱に入ります。 カエルの内部構造により、膀胱が満杯になると筋肉が収縮します。 尿は総排泄腔に入り、そして排出されます。
代謝
かなりゆっくりと進みます。 カエルの体温は周囲の温度にも依存します。 寒い時期には減少し、暖かい時期には増加します。 極度の暑さでは、皮膚からの水分の蒸発により、動物の体温が低下します。 これらは変温動物であるため、寒い天候が始まると活動が鈍くなり、暖かい場所を選びます。 そして冬には完全に冬眠します。
感覚器官
カエルの内臓の構造と機能は、カエルがその生活条件に適応するのに役立ちます。
- カエルは動くことができる上まぶたといわゆる瞬膜を備えており、瞬きすることができます。 目の表面に潤いを与え、付着した汚れを取り除きます。 動物は動いている物体に対してより反応しますが、静止している物体は十分に見えません。
- 補聴器は内耳と中耳で構成されます。 後者は、一方が中咽頭に開いた空洞であり、もう一方は頭の表面に達し、鼓膜によって外部環境から隔離されており、鼓膜はあぶみで内耳に接続されています。 それを通して、音の振動が鼓膜から内耳に伝わります。
- この動物は匂いを頼りにナビゲートするのが非常に得意です。 嗅覚器官は鼻孔を通じて外部環境と通信します。
結論
したがって、カエルの内部構造の特徴は、他の両生類と同様に、感覚器官だけでなく神経系のより複雑な構造にあります。 さらに、肺と 2 つの循環円があります。
超級陸生脊椎動物 - 四足動物
クラス両生類 - アンフィビア
トピック 9. アンフィビバンの廃棄
オブジェクトの体系的な位置
脊椎動物亜門、脊椎動物
クラス両生類、両生類
アヌラン目、アヌラ (Ecaudata)
代表者 - 草のカエル、Rana Temporaria L.
材料と設備
1 人または 2 人の生徒の場合、次のものが必要です。
1. 殺したばかりのカエル。
2. お風呂。
3. メス。
4. 解剖学的ピンセット。
5. 外科用はさみ。
6.準備針-2。
7.ピン10-15本。
8. 延長端を備えたガラス管。
9.脱脂綿。
10. ガーゼナプキン - 2.
生きたカエルを1匹、ガーゼで縛った円筒形の瓶に入れて各テーブルに置きます。
エクササイズ
カエルの外観の特徴を知り、生きているカエルがどのように呼吸しているかを観察してください。 カエルを開いて、主要な臓器系の構造を調べます。 次の図面を作成します。
1. カエルの外観。
2. 口腔。
3. 循環系のスキーム。
4. 内臓の一般的な位置。
5. 泌尿生殖器系は、開いた物体とは異なる性別を持っています。
追加のタスク
1. カエルの皮膚の一部を顕微鏡で観察します。
2. 湿式調製物を使用して、カエルの胎生後発達の特徴 (オタマジャクシ、変態のさまざまな段階) をよく理解します。
外観
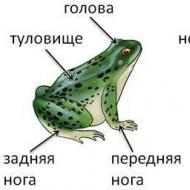
カエルの体は、頭、胴体、前肢、そして長い後肢(ジャンプ運動に適応)に分かれています。 首は外部に表現されていません(図 42)。 前肢は肩、前腕、4本の指で終わる手で構成され、後肢は太もも、下肢、足で構成され、5〜6本の長い指が薄い革のようなひだ、つまり遊泳膜(水泳に適応したもの)でつながっています。 オスの前肢の最初の(内側の)指の付け根には、生殖器疣贅と呼ばれる腫れがあり、交尾中にメスを保持するのに役立ちます。
広くて平らな頭の側面には、大きな凸状の目があり、座りがちな上まぶたとよく動く下まぶたを備えています(死んだカエルのまぶたはピンセットを使用して開閉します。生きているカエルのまぶたの動きを観察してください) )。 銃口の端に近づくと、一対の外部鼻開口部、つまり鼻孔(鼻孔、図42、1)があります。 目の後ろ、口角の上には、半骨化した軟骨輪の上に広がる皮膚の丸い領域があります。 これは鼓膜 (図 42、2) で、中耳腔の入り口を覆っています。 内側から、耳小骨であるあぶみ骨が鼓膜の中心に取り付けられています。
オスのアオガエル(池、Rana esculenta L.および湖、R.ridibunda Pall)の口の隅には、皮膚の薄いひだ、つまり声嚢または共鳴器(図42、3)があり、鳴くときに膨らみます。 (生きている男性の前肢の後ろの体の側面を指で押すと、共鳴器が薄壁の球状の膨らみの形で膨張します)。
米。 42. 雄のイシガエルの出現:
1 - 鼻孔、2 - 鼓膜、3 - 共鳴器、4 - 背側横ひだ、5 - 総排出腔開口部
オスのアカガエル(草ガエル、R. Temporaria L. を含む)では、小さな共鳴器が皮膚の下に隠されています。 鳴くと、腫れて口角の下の皮膚が持ち上げられます。
ラナ属のカエルの体の側面には、皮膚の縦方向の肥厚があります-背側-横方向のひだ(図42、4)。 体の端の上部には、総排出腔の開口部があります(図42、5)。
カエルの体の腹側は明るく、背側は暗く、保護色です。 茶色のカエルでは、黒い縞が目から鼓膜を通って伸びており、目を隠しています。 柔らかく、薄く、粘液腺が豊富な皮膚には鱗がなく、ほぼ全身にある大きな皮下リンパ腔のおかげで非常に動きやすい(体から簡単に引き離せます)。

米。 43. カエルの皮膚の構造図(断面図):
1 - 表皮、2 - 真皮、3 - 皮膚腺の腺細胞、4 - 腺の筋肉被覆、5 - 皮膚腺の排泄管、6 - 色素細胞
魚とは異なり、両生類は体節筋が大幅に減少しており、脊椎に沿った筋肉の比較的弱く発達した部分のみが保存されています。 基本的に、筋肉系は筋肉の部分化の原理に従って構造化されており、筋肉の高度に分化した部分がさまざまな高度に特殊化された機能を実行します。 このタイプの筋肉構造は、地上環境で複雑な動きを実行するタスクに最も適しています。
口腔の構造
口角の関節をハサミで切り、口腔を大きく開けて構造を調べます。 まず第一に、口腔の大きさと口の広い開口部が印象的です。 これは獲物を捕らえやすくするだけでなく、呼吸にも重要です (p. 83)。
均一な小さな歯(図 44、1)が上顎の内側面まで成長します。 下顎には歯がありません。

米。 44. カエルの口腔:
1 - 歯、2 - 舌、3 - 鋤歯のある鋤骨、4 - 蝶穴、5 - 半透明の眼球、6 - 耳管の開口部、7 - 喉頭、8 - 共鳴器の開口部
自由端で二股に分かれた筋肉質の粘着性の舌(図44、2)は、その前端が下顎の前端に取り付けられており、獲物を捕まえるときに口から投げ出すことができます。 口蓋(口腔の屋根)には小さな骨がはっきりと見えます - 小さな鋤骨の歯がその上に座っている鋤骨(鋤骨、図44、3)。 鋤骨の前には、内鼻孔の対の開口部、またはチョアナエがあります(図 44、4)。 針の端をそれらに挿入して、それらが外の鼻の開口部とつながっていることを確認します。 口蓋の中央には、眼球がはっきりと見えます(図44、5)。 目の筋肉が収縮すると、目が口腔内に突き出て、食べ物を食道に押し込むのに役立ちます。 目を上から軽く押して、口腔内にどれだけ深く押し込めるかを確認します。 口腔の深部、顎関節の近くには、中耳腔につながる耳管(チューバ・エウスタキ、図44、6)の開口部があります。 耳管の開口部に挿入された針が鼓膜を通って出ます。
口腔底部の舌の後端の間のフォークには、縦方向のスリットのある小さな隆起が見えます-喉頭(喉頭;図44、7)、対の披裂軟骨によって形成されます。 喉頭のスリットを通って、空気が肺に入ります。 口腔の底部、口角近くに、オスには共鳴器につながる小さな開口部があります(図44、8)。 喉頭裂の後ろで、口腔は気づかないうちに広い食道に入ります。
オープニング
1. カエルの手足を伸ばし、仰向けにして浴槽に置き、腹部の下部の皮膚をピンセットで剥がし、ハサミで切ります(図45)。
2. ハサミの鈍枝を切開部に差し込み、下の筋肉を傷つけないように常に皮膚を上方に引っ張りながら、体の後端から口腔口まで切開します。

米。 45. カエルの解剖の流れ
A - 皮膚を切ります。 B - 体壁を切断します。 B - 本体壁のネジを外します。
1 - 顎間筋、2 - 肩甲帯の筋肉、3 - 腹壁の筋肉、4 - 腹部静脈。 点線 - カットライン
3. 前肢の領域で皮膚を横方向に切開します (図 45、A)。 皮膚フラップを横に回し、ピンで固定します (図 45、B)。 ワックスにピンを斜めに差し込みます。 皮膚を剥がすときは、皮膚が下の筋肉に付着しているのは少数の領域だけであることに注意してください。 残りの空間は皮下リンパ管の空洞によって占められています。
下顎の枝の間には、呼吸機構において重要な役割を果たしている広い顎間筋が見えます (図 45、B、1)。 さらに後方には肩甲帯の筋肉の複合体があり(図45、B、2)、それを強化し、手足の動きを確保します。 準量構造を有する腹筋がはっきりと見えます (図 45、B、3)。 尾のない両生類のここと脊柱の筋肉だけが、筋肉の条件等色性をまだ保存しています。 腹部の正中線に沿って、腹部の静脈(図45、B、4)とそれに流入する腹壁の静脈である暗い帯が見えます。
4. ピンセットで腹部後部の筋肉壁を上に引っ張り、それを切り込み、切り口にハサミの鈍い枝を挿入し、常にそれを使って筋肉壁を持ち上げます(内臓を損傷しないように) 090 に吊り下げ、腹部静脈の側面 (図 45、B) から口腔の始まりまで 3 ~ 4 mm 前方に切ります。 前肢の帯は、その下に心臓とそこから伸びる血管があり、特に注意深く切断されます。 図のように2回目のカットを行います。 45、B、同様に、鼻は腹部の静脈の反対側にあります。
5. ピンセットを使用して、筋肉壁を注意深く側面に引っ張り(必要に応じて、内臓に向かう薄膜をハサミで切り取ります)、ピンで固定します(図 45、B)。 腹部の静脈は筋皮弁の所定の位置に残ります。
重度の出血の場合(解剖手順の指示に注意深く従うことで回避できます)、脱脂綿の塊で作った綿棒を使用して血液を止め、取り除きます(いかなる場合でもプレパラートを水で洗い流さないでください)。 。
内臓の一般的な地形
循環系。 準備の上部では、薄い膜によって形成された心膜腔(心膜腔)内の肺と肝臓の間に、心膜嚢(心膜)があり、心臓があります(図46、47、48)。 ; 時々、開いた後もゆっくりと収縮し続けることがあります。 心臓の上部にある心膜嚢の薄い無色の膜をピンセットで剥がし、心臓を傷つけないように慎重にハサミで切ります。 心臓がバッグから滑り落ちます。 心臓を開かないで、後ろ向きの頂点をピンセットで持ち上げます。 明確な境界のない、黒く薄い壁の静脈洞 (静脈洞) が見えます。これは、2 つの大きな前後大静脈の融合によって形成されます。 心臓の上部には、大きな右心房 (右心房、図 46、1、図 47、2、図 48、1、静脈洞が開口しています) と左心房 (心房洞、完全に開いています) があります。互いに分離されている).図46、2;図47、3 - 右側の標本上;肺静脈がそれに流入します)。 外側では、心房間の境界は非常に弱く表現されます。 心臓の下部のピンクがかった円錐形の最も筋肉の部分が見えます - これは心室です(心室、図46、5、図47、4、図48、3)。 両方の心房は共通の開口部を介して心房と通信します。 この穴の特別な弁(図48、4)により、血液は心房から心室への一方向にのみ流れることができます。

米。 46. カエルの動脈系
動脈血はまばらな陰影で表示され、
混合 - 濃いシェーディング、静脈 - 黒:
1 - 右心房、2 - 左心房、3 - 心室、4 - 円錐動脈、5 - 動脈幹、6 - 皮膚肺動脈、7 - 肺動脈、8 - 大皮動脈、9 - 右大動脈弓、10 - 左大動脈弓、11 - 後頭椎骨動脈、12 - 鎖骨下動脈、13 - 背側大動脈、14 - 腸間膜動脈、15 - 尿生殖器動脈、16 - 総腸骨動脈、17 - 総頚動脈、18 - 内頚動脈、19 - 外頚動脈動脈、20 - 頸動脈、21 - 肺、22 - 肝臓、23 - 胃、24 - 腸、25 - 精巣、26 - 腎臓
動脈円錐(円錐動脈、図 46、4、図 48、5)は心室の右側から伸びています。 したがって、両生類の心臓は 3 室 (2 つの心房と 1 つの心室) ですが、静脈洞、2 つの心房、心室、円錐動脈という 5 つのセクションで構成されています。 円錐動脈は 3 対の動脈弓を生じます。 各アーチは独立した開口部を持って動脈円錐から離れています。 左側と右側の 3 つのアーチはすべて、まず共通の膜で囲まれた共通の動脈幹 (動脈幹、図 46、5、図 48、6) と一体になっているため、動脈円錐は 2 つの大きな幹にのみ分割されます。
しかし、この幹を針で持ち上げると、別々ではあるが隣接した血管で構成されていることがはっきりとわかります。 これらの血管(動脈円錐筋からの起源の順)は次のとおりです。
1. 動脈円錐の背側セクションから最初に離れるのは、一対の(右と左)皮膚肺動脈(肺動脈、図46、6;図48、7)です - 鰓動脈のIV対の相同体魚のアーチ。 すぐに、各皮膚肺動脈は肺動脈 (肺動脈、図 46、7) に分かれ、肺の端に沿って肺の頂点まで通過し、大きな皮動脈 (皮膚大動脈、図 46、8) に分かれます。 )、背側体表面の皮膚で分岐します。
2. 皮膚肺動脈のすぐ後ろにありますが、動脈円錐の腹部からは、一対の大動脈弓が伸びています(大動脈弓、図46、9、10、図48、8)。 それらは、鰓動脈弓の 2 番目のペアと相同です。 各大動脈弓は上方(体の背側表面に向かって)および側方に湾曲し、後頭椎骨(後頭椎骨動脈、図46、11)と鎖骨下動脈(鎖骨下動脈、図46、12)を分離し、前肢に血液を供給します。血液)動脈。 次に、大動脈弓は脊柱の下(胃の後部のレベル)で互いに結合し、対になっていない背側大動脈(背側大動脈、図46、13)に入ります。 強力な腸腸間膜動脈 (arteria coeliaco-mesenterica; 図 46、14) が背側大動脈から出発しています。 それは腸間膜のひだに沿って通過し、血液を胃、腸、肝臓、脾臓に運びます。 逆向きに走る背側大動脈(そこから出発した腸間膜動脈よりも細い幹のように見える)は、腎臓と生殖器に細い動脈を数本出しています。 腎臓の後端のレベルで、背側大動脈は 2 つの総腸骨動脈 (総腸骨動脈、図 46、16) に分岐し、その枝は体の後部と後肢に血液を供給します。
3. 頭部に血液を供給する頸動脈弓 (頸動脈) は、大動脈弓に沿って、また総頸動脈 (総頸動脈、図 46、17; 図 46、17) の形で動脈円錐の腹部からも続きます。 48、9)。 総動脈幹を出たほぼ直後に、各頸動脈弓は外頸動脈 (外頸動脈、図 46、19) と内頸動脈 (内頸動脈、図 46、18) 動脈に分かれます。 それらの分岐箇所、内動脈の基部には頸動脈、または頸動脈「腺」(頸動脈腺、図 46、20、図 48、10) があり、頸動脈内の血圧を明らかに調節しています。 。

米。 47. カエルの静脈系
静脈血は黒で示され、動脈血は網掛けで示され、混合血液は点で示されます。
1 - 静脈洞、2 - 右心房、3 - 左心房、4 - 心室、5 - 外頸静脈、6 - 内頸静脈、7 - 大皮静脈、8 - 上腕静脈、9 - 鎖骨下静脈、10 - 右前大静脈、11 - 左前大静脈、12 - 大腿静脈、13 - 坐骨静脈、14 - 総腸骨静脈または腎門脈、15 - 腹部静脈、16 - 肝門脈、17 - 遠心性腎静脈、18 - 後大静脈、19 - 肝静脈、20 - 肺静脈、21 - 肺、22 - 肝臓、23 - 胃、24 - 腸、25 - 精巣、26 - 腎臓
頭部からの静脈血は、外頸静脈と内頸静脈を通過します(外頸静脈および内頸静脈;図 47、5、6)。 皮膚で酸化された動脈血は、強力な太い皮静脈(皮大静脈、図47、7)を流れ、そこに前肢からの静脈血を運ぶ上腕静脈(上腕静脈、図47、8)が流れます。 。 皮膚静脈と上腕静脈は鎖骨下静脈に合流します(鎖骨下静脈、図 47、9)。 ほぼ即座に、両側の鎖骨下静脈が外頸静脈と内頸静脈と合流し、右 (右大静脈前腕部、図 47、10) と左 (大静脈前洞、図 47、11) 前大静脈を形成します。 。 動脈血と混合した静脈血を運ぶ(大皮静脈を通って入る)両方の前大静脈は、静脈洞に流れ込みます。
後肢と骨盤領域から、静脈血がいくつかの静脈を通って流れます。 それらの中で最大のものは大腿静脈(大腿静脈、図 47、12)と坐骨静脈(坐骨静脈、図 47、13)であり、両側で合流して対の総腸骨静脈、または腎門脈(静脈)を形成します。腎門;図 47、14)、腎臓に到達し、そこで毛細血管のネットワーク (腎臓の門脈系) に分裂します。
幹は左右の大腿静脈から出発し、腹部静脈に互いに合流します(腹部静脈、図47、15)。 それは体の腹壁に沿って通過し、膀胱と筋肉から血液を集め、胸骨の後端近くで腹腔に突入し、肝臓に進み、そこで毛細血管に分裂します。 腸、胃、食道のすべての部分からの静脈血は、静脈系を通って肝臓の大きな門脈 (肝門部静脈; 図 47、16) に集められ、肝臓に入り、そこで毛細血管に分裂します。 したがって、両生類では、肝臓の門脈系は肝臓自体の門脈と腹部静脈の2つの静脈によって形成されています。
腎臓の毛細血管を通過する静脈血は、いくつかの遠心性腎静脈 (腎静脈、図 47、17) に集まり、対になっていない後大静脈 (後大静脈、図 47、18) に合流します。 生殖腺から血液を運ぶ静脈がそこに流れ込みます。 後大静脈はすぐに肝臓の中央部に入り、肝臓を貫通します(そこからの血液は肝臓に入りません!)。 肝臓の出口で、後大静脈は2本の短い肝静脈(肝静脈;図47、19;肝臓のすべての部分から血液を集める)を受け取り、静脈洞に流れます。
肺からの動脈血は肺静脈 (肺静脈、図 47、20) を通過し、合流して左心房に流れ込みます。 それらの合流点は左前大静脈で覆われています。
両生類では、小さい血液循環(心室 - 肺 - 左心房)と大きい血液循環(心室 - 全身 - 静脈洞 - 右心房)の 2 つの血液循環が明確に表現されます。 ただし、心室が 1 つであるため、これらの円はまだ完全に分離されておらず、血液の一部が心室内で混ざります。 両生類の活動状態では、肺と皮膚の両方で血液の酸素飽和が起こります。 したがって、右心房には、静脈血(全身の静脈から静脈洞に集められる)と動脈血(大きな皮静脈によってもたらされる)が混合されています。 左心房では、血液は動脈血です(肺から肺静脈を通って来た)。 心房も同時に収縮し、血液が心室に入ります。 筋肉の成長が強力に発達しているため、心室の空洞は、いわば多数の部屋に分割されており(図48)、血液の混合を妨げます。 したがって、心室の右部分には動脈血と混合した静脈血(右心房と同じ組成)があり、心室の左部分には動脈血(左心房と同じ)があり、真ん中部分は血が混ざっています。

米。 48. 開いたカエルの心の図式:
1 - 右心房、2 - 左心房、3 - 心室、4 - 弁、
両心房から心室につながる共通の開口部を閉じる、
5 - 円錐動脈幹、6 - 総動脈幹、7 - 肺皮動脈、8 - 大動脈弓、9 - 総頸動脈、10 - 頸動脈、11 - 円錐動脈らせん弁
心室から動脈幹への血液の流れはまだ十分に研究されていません。 このプロセスは次のように簡略化できます。 心室が収縮すると、より多くの静脈血がまず動脈円錐形筋に流入します(その起源が心室の右側からであるため)。 それは開いた開口部を通って直ちに皮膚肺動脈を満たし(残りの動脈弓の開口部は動脈円錐の螺旋弁によって閉じられている)、肺および皮膚に入り酸化される。 継続的な心室収縮によって肺皮動脈が満たされた後、円錐動脈内の圧力が増加します。 らせん弁が動き、大動脈弓の開口部が開きます。 混合血液は心室の中央部からそれらに突入し、大動脈弓の枝と背側大動脈の枝に沿って体全体に広がります。
心室の最大収縮中に心室の左側から円錐動脈に出る動脈血は、すでに血液で満たされているため、皮膚肺動脈および大動脈弓に入ることができません。 螺旋弁が最大に移動し、頸動脈の口が解放されます。 それらを通って、動脈血は頭部(脳および感覚器官を含む)に送られます。
尾のない両生類において、肺呼吸が長期間停止しているとき(たとえば、皮膚の表面でのみ呼吸が行われる貯水池の底で越冬しているとき)における血流の分離のメカニズムはまだ解明されていない。 尾状両生類では、円錐動脈の螺旋弁が十分に発達していないため、より多くの混合血液がすべての動脈弓に流入します。
呼吸器系。 呼吸器系には、経路と肺の両方が含まれます。 喉頭裂は、いくつかの軟骨によって口腔から区切られ、小さな空洞である喉頭(喉頭)に通じています。 喉頭裂は、喉頭の特別な筋肉の収縮によって開閉できます。 披裂軟骨の凹面の内側には声帯、つまり喉頭の粘膜のひだがあります。 空気が喉頭を通過することによってこれらの靭帯が振動すると、共鳴器によって増幅された音(きしみ音)が発生します。 喉頭腔からの 2 つの小さな開口部は、対の肺に直接つながっています。

米。 49. メスのカエルの内臓の一般的な配置:
1 - 右心房、2 - 左心房、3 - 心室、4 - 円錐動脈、5 - 肺、6 - 食道、7 - 胃、8 - 胃の幽門部、9 - 十二指腸、10 - 膵臓、11。 - 小腸、12 - 直腸、13 - 総排出腔領域、14 - 肝臓、15 - 胆嚢、16 - 胆管、17 - 腸間膜、18 - 脾臓、19 - 腎臓、20 - 尿管、21 - 膀胱、22 - 卵巣、23 - 卵管
(左卵巣と卵管は図には示されていません)
心臓の側面にある肺(肺、図49、5)は、弾性壁を備えた薄壁の嚢であり、外側には細胞構造を持っています(肺をよりよく検査するには、ガラス管を挿入してわずかに膨らませる必要があります)細い端を喉頭スリットに挿入します)。
細胞性は、肺の壁の内側にある小さな増殖物(隔壁)によって引き起こされ、これにより、その内面がわずかに増加します。 しかし、両生類の肺の内面の総面積は小さく、通常は皮膚の表面よりも若干小さい(いくつかの種ではわずかに大きい)ことさえあります。 (哺乳類では、肺の内面は皮膚の表面の 60 ~ 100 倍も大きくなります。)
実際の陸生脊椎動物(爬虫類、鳥類、哺乳類)では、空気は主に胸部(肋骨を介した胸骨と脊柱の接続部)の容積を変化させることによって肺に吸い込まれます。 両生類には胸がありません(肋骨がないか、または非常に発達が不十分です)、そしてその呼吸メカニズムは非常に独特です。 生きているカエルでは、口の底がリズミカルに上がったり下がったりするのがはっきりとわかります。 鼻孔の外側の開口部は、異なるリズムで開閉します。

米。 50. カエルの呼吸メカニズムの図解
I - 口腔が拡張し、開いた鼻孔から空気が入ります。 II - 鼻孔が閉じ、喉頭スリットが開き、肺から出た空気が口腔内で大気と混合します。 III - 鼻孔が閉じられ、口腔が収縮し、混合された空気が肺に送り込まれます。 IV - 喉頭裂が閉じられ、口腔の底が口蓋に押し付けられ、残った空気が開いた鼻孔から押し出されます: 1 - 鼻孔の外側の開口部、2 - 鼻孔の内部開口部 (チョアナ)、3 - 口腔、4 - 口腔底、5 - 喉頭間隙、6 - 肺、7 - 食道
口腔の底が下がると、口腔の体積が大幅に増加し、空気が鼻道(開いた外鼻孔と鼻腔)を通って口腔内に吸い込まれます。 この時点では、喉頭裂は閉じています (図 50、I)。 次に、外鼻孔が閉じます(これは鼻孔の特別な筋肉の作用によって起こります。位置を変える上顎前骨の突起も閉じるのに役立ちます)、同時に喉頭裂が開きます。 空気は肺から(内臓からの圧力と腹壁の筋肉の収縮を受けて)膨張し続ける口腔に入り、そこにある大気と混合します(図50、II)。
次に、口腔の底部が口蓋まで徐々に上昇し始め、口腔から混合された空気が肺に押し込まれます(図50、III)。 次に喉頭裂が閉じ、口腔の底が口蓋に押し付けられ、残った混合空気が開いた鼻孔から押し出されます(図50、IV)。 その後、吸入の最初の段階が再び始まります。
不規則な呼吸運動の合間に、口腔底は鼻孔が開き、喉頭スリットが閉じた状態で小さな振幅の振動を起こします。 同時に、口腔内の空気が更新され、口腔粘膜の毛細血管内の血液が酸素で飽和されます。
消化器系。 消化管は口腔から始まり総排出腔で終わります。 喉頭裂の後ろの口腔 (75 ページを参照、図 44 を参照) から、短くて容易に伸長可能な食道 (食道、図 49、6) が広い開口部で始まり、上方の体腔の背側に沿って通過します。心臓、肺、肝臓。 肝臓の右葉(開口部から)を左に回すと、その後ろの部分がはっきりと見えます。
食道は胃(胃;図49、7)に流れ込み、リング状の狭窄部によって胃から分離されます。 胃はわずかに湾曲しており、食道よりも厚い筋肉の壁を持っています。 胃の狭くなった後端(幽門、図49、8)は、小腸の最初の部分である十二指腸(十二指腸、図49、9)からほとんど目立たない環状の狭窄によって分離されており、十二指腸は胃と平行に伸びています。お腹が前に出ます。 胃と十二指腸の間の腸間膜には、膵臓がゆるやかな黄色がかった紐の形で存在します(膵臓、図49、10)。 明確な境界がなく、十二指腸は小腸(回腸、図49、11)に入ります。小腸はわずかに小さな直径を持ち、体腔の右側にいくつかのループの形で位置しています(開口部から上に向かって)。左側)。 小腸は短くて広い直腸(直腸、図49、12)に入り、薄くなり、背側に開口部のある総排出腔(総排出腔)を形成します(図42、5を参照)。
肝臓(hepar、図49、14)は、大きくてコンパクトな3葉の器官であり、心臓のすぐ後ろにあります。 その小さな中葉の下面には、丸い緑がかった黒色の胆嚢(vesica felea、図49、15)があります-肝臓によって分泌された胆汁が蓄積する貯留槽です。 肝管は肝臓の右葉と左葉の背側表面に沿って通り(注意深く検査した場合にのみ見ることができます)、胆嚢管(特別な準備がなければ見ることができません)と一緒に総胆管(総胆管; 胆管)に合流します。図49、16)。
密なコードの形をした胆管は膵臓の組織を通過し(図49、10)、そこからいくつかの小さな短い管を受け、十二指腸の最初の部分に流れ込みます。 肝臓、胃および腸全体は、体腔の背側表面から薄い半透明の折り畳まれたフィルム、つまり腸間膜上に吊り下げられています(腸間膜;図49、17)。
食道の機能は食べ物を胃に運ぶことです。 ここで、食べ物は筋肉の壁の圧力によって押しつぶされ、胃の腺から分泌される消化酵素に浸されます。 小腸では、食物塊に肝臓と膵臓から胆管を通って入ってくる酵素が含浸され、消化され、腸壁から吸収されます。 直腸では、水分が吸収され、便が形成され、総排出腔を通って放出されます。
小腸の腸間膜には(開いたカエルの上、通常は胃と直腸の間に)、細網内皮系の非常に重要な器官である小さな丸い脾臓(リアン、図49、18)があります。 脾臓では、形成された血液要素(赤血球、リンパ球)の形成が起こり、血液に入る細菌の食作用などが起こります。脾臓は、必要に応じて血液の貯蔵庫としても機能します(失血、長時間の激しい動きなど)。 )血流に放出されます。
泌尿器系。 一対のコンパクトな細長い楕円形の中腎(または幹)腎臓(腎、図 49、19、図 51、1、図 52、1)は、体腔の後部の脊柱の側面に位置しています。 。 各腎臓の外縁に沿って、薄い灰色がかったコード、つまりウォルフィ管(ウォルフィ管、図 49、20、図 51、2、図 52、2)、つまり中腎腎臓の尿管が走っています。 左右の腎臓のウォルフ管は総排出腔の背側部分で独立した開口部を持って開いています(図51、4;図52、4)。 総排泄腔の腹壁にある不対の開口部は、広範な二葉の薄壁膀胱につながっています(膀胱尿路、図 49、21、図 51、5、図 52、5)。 ピペットを使って総排出腔を通って膀胱の開口部に水を導入すると、膀胱壁の強い伸展性を簡単に確認できます。 ウォルフ管(尿管)を通って総排出腔に入った尿は、総排出腔の底に流れて膀胱に入ります(図53)。 その壁にはかなり密な毛細管ネットワークがあり、尿からの水分が確実に吸収されます。 尿はより濃縮され、膀胱壁の収縮が再び総排出腔内に排出され、総排出腔から排出されます。

米。 51. 雄のカエルの泌尿生殖器系:
1 - 腎臓、2 - 尿管(精管としても知られる)、3 - 総排出腔、4 - 泌尿生殖器の開口部、5 - 膀胱、6 - 膀胱の開口部、7 - 精巣、8 - 精細管、9 - 精嚢、10 - 脂肪体、11 - 副腎
これらの特徴は、男女両方の排泄システムの特徴です。
各腎臓の前端では、生殖腺と同じ腸間膜に、指の形をしたオレンジ色の脂肪体が位置しています(図51、10;図52、10)-生殖細胞の形成のための栄養素の貯蔵庫です。 各腎臓の表面に沿って、狭く、時にはかすかに目立つ黄色がかった帯-副腎(副腎体、図51、11;図52、11)-内分泌腺が伸びています。
精巣(精巣;図51、7) - 対になった、丸く、黄色がかった色または茶色がかった色で、腎臓の前端近くの脂肪体とともに腸間膜に浮遊しています。 ピンセットで精巣を注意深く引っ張ると、精巣から伸びる細い白っぽい糸が腸間膜 - 精細管(精管;図51、8)で目立ち、腎臓の前部に流れ込みます。
精細管は尿細管に開口しており、したがって両生類の腎臓の前部は機能的に精巣上体として機能し、雄の両生類のウォルフ管は同時に尿管と精管の両方として機能します。 繁殖期(4月、5月)には、芽から出た直後にウォルフ管の壁にポケット状の拡張がはっきりと見えます-精嚢(vesica seminalis;図51、9)。 それらは精液の貯蔵庫として機能します。 繁殖期以外では、精嚢のサイズは減少しますが、依然として目で見ることができます。

米。 52. メスのカエルの泌尿生殖器系:
1 - 腎臓、2 - 尿管、3 - 総排泄腔、4 - 排尿口、5 - 膀胱、6 - 膀胱開口部、7 - 左卵巣(右卵巣は図には示されていない)、8 - 卵管、9 - 卵管漏斗、10 - 脂肪体(右側の脂肪体は示されていません)、11 - 副腎、12 - 生殖口(卵管の開口部)
一対の卵巣(卵巣、図52、7)は腸間膜上に(脂肪体とともに)吊り下げられた薄壁の嚢であり、季節に応じて体腔の多かれ少なかれ重要な部分を満たしています。 卵巣を埋める色素のついた卵が、卵巣の壁を通してはっきりと見えます。 女性の生殖管は一対の卵管、つまりミュラー管(卵管、またはミューレン管;図52、8)であり、体腔の側面の短い腸間膜に吊り下げられています。 卵管の長さは季節によって大きく異なります。 繁殖期の春には特に強く曲がりくねって伸びます(体長の6〜8倍を超えます)。 各卵管の前端は、拡張された開口部、つまり卵管の漏斗を備えた体腔(心臓の隣)に開口しています(図52、9)。 卵管の下部(子宮部分と呼ばれることが多い)は幅が広くなります。 各卵管は独立した開口部で総排泄腔に通じています(図 52、12)。
米。 53. メスのカエルの総排出腔の図:
1 - 総排出腔の外部開口部、2 - 総排出腔の空洞。 3 - 直腸、4 - 膀胱、5 - 尿管、6 - 卵管、7 - 体壁
卵子が成熟すると、周囲の卵胞膜が破裂し、卵子が体腔内に押し込まれます。 ここで、それは卵管漏斗によって拾われ、産卵の開始時に急激に増加し、その壁の蠕動運動のおかげで卵管に沿って移動します。 この場合、卵管の壁にある腺の分泌物が、卵の周りに透明なゼラチン状の卵殻を形成します。 卵管の下部(子宮)では、完全に形成された卵(卵子)が、産卵の準備ができた塊にまとめられます。
したがって、両生類(典型的な無羊症)では、軟骨魚と同様に、成体になっても中腎型の腎臓が機能します。 ヴォルフの雄では、この管が尿管と精管の両方の役割を果たし、精子が体腔に落ちることはなく、ミュラー管は縮小しています。 ウォルフ管のメスでは、管は尿管としてのみ機能し、ミュラー管は卵管として機能します。 成熟した卵子は体腔に落ち、漏斗を通って卵管に入ります。
末梢神経系。 すべての器官系をスケッチした後、解剖したカエルの内部を取り除くと、脊柱から伸びている密な白いコード、つまり脊髄神経がはっきりと見えるようになります。 脊柱の側面には、リンパ管を通って内耳の被膜の膜迷路の空洞につながっている白い「石灰質の嚢」が見えます。 これらの嚢の機能的重要性はまだ明らかになっていません。
カエルの胎生後の発達。 湿った標本でカエルの幼虫、オタマジャクシの発育段階を調べます。 卵から孵化した直後のオタマジャクシには、外えらと小さな尾があります。 オタマジャクシが成長するにつれて、革のようなひだ (「鰓蓋」) が発達して外鰓を覆い、尾が成長します。 変態は徐々に進行します。四肢の基礎が現れ(後肢と同時に形成される前肢の基礎は、外えらを覆う革のようなひだで覆われているため見えません)、成長と形成が進みます。 。 尾は徐々に溶け始めます。 こうした外部の変化と同時に、消化器系、呼吸器系、循環器系といった内臓の再構築が起こります。 変態の結果、水生幼生は水の外で生きることができる小さなカエルに変わります。
カエルの心臓は、食道の下の体の腹側、咽頭からそれほど遠くない場所に位置し、薄い膜、漿液膜、心膜(心膜)で覆われた心膜腔に囲まれています。 心臓自体は、背側に位置する静脈洞、緻密な筋肉の心室(図 2、3)、壁が薄い 2 つの心房、および動脈円錐、または大動脈の円錐(図 2、4)で構成されています。 静脈洞は右心房に開口しています (図 2、9)。 左側に肺静脈(図2、10)。 心房は完全な中隔によって分割されています(図2、7)。 彼らは心を開きます
米。 1. カエルの血液循環図。
1-内頸動脈。 2-鎖骨下静脈。 3-皮動脈。 4-肺動脈。 5-大動脈; 6-肺静脈。 7内臓動脈。 8-皮静脈。 9後大静脈。 10 腎臓の門脈。 11-腸骨静脈。 12-坐骨静脈。 13-腸骨動脈。 14-腹部静脈。 肝臓の15番目の門脈。 16-肝静脈。 前肢から17本の静脈。 18-前肢への動脈。 19 - 前大静脈。 20総頸動脈。 21-無名静脈。 22-外頸静脈。 23-外頸動脈。
1 つの共通の開口部を備えた総心室に挿入され、一対の弁で保護されています。 大動脈の円錐は心室の基部の右側から生じます。 円錐形の原点には 3 つの小さなバルブがあります。 縦方向の羽根状のバルブがコーンに沿って伸びています (図 2、5)。 円錐体自体は、直径を変えることなく大動脈球に入り、右と左の2つの枝が生じます。 各ブランチは 3 つの容器に分かれています。 上部は頸動脈幹を表し(図 2、11)、中央は全身大動脈弓を表し(図 2、12)、下部は肺皮膚幹を表します(図 2、13)。 。
頸動脈幹の基部には、血管叢からなる頸動脈腺の小さな腫れがあります。 全身幹、または大動脈弓は咽頭の周りで曲がり、その下でつながって背側大動脈を形成し(図1、5)、そこから動脈血管がすべての内臓、腸、生殖器、腎臓に伸びています(図1、5)。 2、7)。 最後に、肺皮幹は肺に向かう肺動脈と皮膚に向かう皮下動脈の2つの枝に分かれています(図1、3、4)。
記載されている主要動脈血管の構造をオタマジャクシの構造と比較すると、成体のカエルでは最初の大動脈弓が背側大動脈との接続を失い、頸動脈の幹に変わっていることがはっきりとわかります。 2番目の弓は厚くなり、背側大動脈との接続を維持して全身幹になります。 3番目の弓は完全に消失します(尾状両生類の構造とは異なります)。4番目の弓は肺と皮膚に枝を送り、背側大動脈から分離します。

図2。 カエルの開いた心臓(腹側から)。
1 - 左心房。 第2右心房。 3心室。 4動脈円錐。 5 ブレード形状のコーンバルブ。 6つの円錐の中央のパーティション。 7 - 心房の間の中隔。 心房と心室の間の 8 弁。 9-右心房への静脈洞の開口部。 10-左心房への肺静脈の開口部。 大動脈弓の頸動脈の第11管。 大動脈弓の12の共通システムチャネル。 13-肺皮管。 心室の14の側室。
末梢の分布の最終部分に近づいている動脈
毛髪または毛細血管網に分解され、小さな静脈が形成されます。 互いに接続して、心臓につながる大きな静脈血管を形成します。 心臓に直接流れる最大の静脈は、4 本の主要な血管で構成されています。 総肺静脈 (右肺静脈と左肺静脈で構成される総肺静脈) は左心房に流れ込みます (図 3、21)。前述したように、血液は心臓から肺枝を通って肺に入ります。肺皮動脈。肺の壁で毛細血管に分裂します。
肺には酸素が豊富な空気が存在するため、二酸化炭素が静脈血に放出され、血液は酸素で飽和されます。 肺静脈は酸素が豊富な血液を供給します。 示されているように、それは左心房に向けられています。 肺と心臓の間で起こる血液循環は肺循環と呼ばれます。
3 つの大きな静脈血管が静脈洞または洞に流れ込みます。左右の上大静脈 (上大静脈、右大静脈、図 3.1)、下大静脈 (下大静脈、図 3.9) 。 各上大静脈は、外頸静脈および内頸静脈 (図 3、2、5) と、上腕静脈 (図 3、7) が入る鎖骨下静脈 (図 3、6) で構成されています。大きな皮静脈 (図 3、8)。

米。 3、カエルの静脈系の図。
1 - 上大静脈(右)。 2-外頸静脈。 3-無名静脈。 4-肩甲下静脈。 5-内頸静脈。 6-鎖骨下静脈。 7-上腕静脈。 8-大皮静脈。 9-下大静脈。 10-肝(遠心性)静脈。 肝臓の第11門脈。 12-腎臓の遠心性静脈。 13および14は外腸骨静脈。 15腸骨横静脈。 16 - 坐骨静脈。 17-大腿静脈。 18-腹部静脈。 19 - 背腰静脈。 心臓球の後部20静脈。 21-肺静脈(右)。 22 - 肺(左)。 23 - 卵巣。 24本の腸管(セグメント)。 25-卵管(セグメント); 26肝臓(一部除去)。
体の後部から心臓までの血液経路は、魚について説明されているものとは大きく異なります。 魚の枢静脈はカエルでは下大静脈に置き換えられています(図3、9)。 後肢から、静脈血は大腿静脈 (大腿静脈、図 3.17) を通って運び出され、体腔内で背側と腹側の 2 つの枝に分かれます。 背静脈は腸骨静脈で構成され (図 3、13、14、15)、坐骨静脈も同じ系に流れ込みます (図 3、16)。 腎門脈とも呼ばれる総腸骨静脈は腎臓に近づき、そこで毛細血管のネットワークに分かれ、腎門脈系を形成します。
耳介枝は骨盤静脈で構成され、重要な腹部静脈と合流します(図 3、18)。 それは体の腹壁に沿って胸骨のレベルまで走り、そこで2つの枝に分かれて肝臓の物質に入り、そこで毛細血管に分解されます。 腸から血液を運ぶ肝口腔静脈(図 3、11)も肝臓に入り、毛細血管網を形成します。 血液は腎臓から腎静脈を通って後大静脈または下大静脈に流れます。 後者は肝静脈を含む肝臓葉の間の切欠きを通って静脈洞に流れ込みます。
大動脈弓を通って静脈を通って静脈洞に戻る血液の経路は、体循環と呼ばれます。
次に、カエルの心臓内で、それに隣接する主要な血管に血液がどのように分布しているかを見てみましょう。
二酸化炭素を多く含む静脈血が大静脈を通って静脈洞(副鼻腔)に流れ込むことはすでに見てきました。 副鼻腔の収縮(収縮)により、副鼻腔と心房を接続する静脈開口部を通って血液が右心房に押し出されます。 同時に、酸素が豊富な血液 (いわゆる「動脈」血) が肺静脈を通って左心房に流入します。 心房の同時収縮(収縮期)により、動脈血(酸素が豊富)と静脈血(二酸化炭素が豊富)が総心室の空洞に流れ込みます。 心房が拡張し始めるとき(拡張期中)および心室収縮期には、胃前部の開口部が 2 つの弁によって閉じられます。
この時点で、心室と心房間の通信は完全に遮断されます。 静脈血は心室の右半分に入り、動脈血は左半分に入ります。 心室の主室では、それらの部分的な混合が起こります。 これは、優れた脊椎動物と比較した両生類の血液循環の不完全さです。 2 つの血流の完全な混合は、2 つの状況によって妨げられます。1) 血液の主要な塊は、心室の下部に位置し、不完全な隔壁によって分離されている、心臓の心室のいわゆる副室に入ります。 2) 心室収縮は非常に速く、これも血流の混合を妨げます。

図4.
I-嗅覚神経。 IV 滑車神経。 VII-顔面神経。 IX-X 舌咽神経および迷走神経、腹側から 6-脳: 1-下垂体。 2 脳漏斗; 3-ビジュアル。 キアズマ; II-視神経。 III - 動眼神経。 V-三叉神経; VI - 神経を外転します。 VII-顔面神経。 VIII - 聴覚神経。 IX-X - 舌咽神経と迷走神経。 12正中裂。 その他の名称は図の通りです。 A. V-側面からの脳: 1-下垂体; 2-大脳、漏斗。 3-視覚視交叉; 4つの視葉。 5 - 小脳。 脳の8半球。 9-嗅葉。 10-秒脊髄神経(舌下神経)。 I-嗅覚神経。 II-視神経。 P1 - 動眼神経。 IV 滑車神経。 VI は神経を外転します。 IX-X - 舌咽神経と迷走神経。
ある非常に短い瞬間に、心臓の心室の左側には動脈血、右側には静脈、中央には動脈血が混合されます。 収縮期には、房胃弁が閉じ、血液が心室基部の右側にある大動脈に流れ込みます。 まず、収縮期の開始時に、心室の右側に蓄積された静脈血が大動脈に流入することは明らかです。 この血液は、血流に対する抵抗が最も少ない大動脈の最も短い肺皮膚幹に沿って流れます。 心室収縮期の第 2 段階では、動脈円錐の壁が収縮し、ブレード状の弁が左に動き、円錐の肺部分が閉じて大動脈幹が開きます。 動脈と静脈の混合血液が彼らに流れ込みます。 心室収縮期の第 3 段階では、肺皮膚幹はブレード型の弁によって閉じられたままですが、大動脈管では以前の充填により、新しい血流に対する抵抗が増加します。 頸動脈幹への血液の最後の純粋な動脈部分の自由な経路が残っています。 毛細血管を備えたいわゆる「眠い」腺は、もはや抵抗することができません。
したがって、カエルの頭には純粋な動脈血の流れが供給されます。 心室拡張期の間、血液は心臓に戻ることができません。
これは半月弁によって防止されます (上記を参照)。

心室に中隔がないにもかかわらず、前述の複雑な構造のおかげで血流の連続的な分布が達成されます。
これは、弁の動作メカニズムのほか、球、大動脈、および心室内の追加の部屋の存在から伸びる 3 つの幹のさまざまな程度の抵抗によるものです。 純粋な静脈血は酸化のために肺皮膚幹に入り、全身幹は混合血液を受け取り、純粋な動脈血は(頸動脈を介して)脳に供給されます。
神経系。 カエルの脳
脳の構造は次のような特徴があります: 1) 中央面で融合した大きな嗅葉 (図 4、9)。 2)かなり大きな前脳。これは魚のものよりも比較的大きい(図4、8)。 3)かなりよく発達した間脳。 4)中脳の大きな視葉(図4、4)。 5)非常に小さな小脳(図4、5)。
カエルの血液循環に関する記事
さまざまな文化におけるカエルの象徴性
魔法では、カエルは月の生き物とみなされ、変容を象徴します。
 その気取らない外観にもかかわらず、この水陸両用の生き物は魔法と魔術に満ちています。 ケルトの伝統では、地球の主と癒しの水の意味があります。
その気取らない外観にもかかわらず、この水陸両用の生き物は魔法と魔術に満ちています。 ケルトの伝統では、地球の主と癒しの水の意味があります。
キリスト教徒にとって、カエルには再生と罪という二重の意味があります。 彼女は貪欲、暴食、そして嫉妬の持ち主だと信じられています。 エジプト人はカエルに対してより慈悲深い態度をとります。これは、この両生類が豊饒、豊かさ、長寿の力を象徴しているためです。 そして、カエルの恋愛ゲームは、ギリシャ人をアフロディーテ自身との神聖なつながりについての結論に導きました。
風水の豊かさのシンボル
中国ではカエルやヒキガエルに対して特別な態度をとります。 それらは富と幸運、豊饒と豊かさに関連しています。 中国のヒキガエルは 3 本の足があり、口にコインをくわえています。 もちろん、三本足のヒキガエルは金で作られている必要がありますが、模造金も金持ちになるのに役立ちます。
風水のカエルのお守りは、富の流れを呼び込むために適切な場所になければなりません。 通常、部屋の南東にある富と幸運の分野に置かれます。
中国のお金のヒキガエルを配置するためのルールは次のとおりです。
- お金のヒキガエルはリビングルームに置くのが良いでしょう。
- ヒキガエルに最も適した場所はパワーアングル(ドアから左斜め)です。
- ヒキガエルは視覚的に家に飛び込むように見えるように立つ必要があります。
- お守りは暗闇の中で隅っこに埃をかぶっているべきではありません。そうしないと役に立ちません。
- ヒキガエルは水と関係があるという事実にもかかわらず、バスルームに置くべきではありませんが、家の小さな噴水に置くのはまさに問題です。
お守りの効果を十分に発揮するには、ヒキガエルを水浴びさせる必要があります。 2週間に1回シャワーで洗うだけで十分ですが、それ以上の頻度で洗ってください。 お金を受け取るたびに、自分の言葉でヒキガエルに感謝しましょう。 お守りの下にお札や小銭を入れることができます。
その他のお守りの種類とその意味
中国のお金のヒキガエルに加えて、月のカエルの魔術師を描いた他のお守りもあります。 これらの工芸品は、私たちによく知られている 4 本足のカエルを表現しており、さまざまな素材で作られています。
金でできたカエルは男性の仕事運を引き寄せ、銀でできたカエルは女性を引き寄せます。
足や口に指輪のあるカエルは家族経営に有利です。
ローズクォーツで作られたカエルは、文化人や芸能関係者にも重宝されるでしょう。
 マラカイトのアーティファクトは、嫉妬深い人々からあなたを守り、あらゆる努力に成功をもたらします。 たった 1 つのお守りだけでこれを行うことはできません。オフィスや書斎にいくつかのマラカイト ワウを置きましょう。 隠れて嫉妬している人にマラカイトカエルを与えると、彼はあなたに対するこの感情を取り除くことができます。
マラカイトのアーティファクトは、嫉妬深い人々からあなたを守り、あらゆる努力に成功をもたらします。 たった 1 つのお守りだけでこれを行うことはできません。オフィスや書斎にいくつかのマラカイト ワウを置きましょう。 隠れて嫉妬している人にマラカイトカエルを与えると、彼はあなたに対するこの感情を取り除くことができます。
オニキスのお守りは、競争相手や敵に対する怒りを鎮め、報われない愛から心を解放するのに役立ちます。 元愛する人をすぐに忘れるには、悲しい記憶がなくなるまで 15 分間、お守りを心に当ててください。
ロッククリスタルのお守りは、新しい環境に素早く適応するのに役立ちます。 別の地域に旅行したり、住む場所を変えたりするときに便利です。
透明な石で作られたお守りは、コミュニケーションにおいて妥協を求め、繊細になる能力を人に与えます。 論争や議論では、そのようなカエルは単に不可欠なアシスタントです。 手で触ったり、机の上に置いたりすれば、衝突を避けることができます。
琥珀色のカエルは女性の美しさに欠かせないものです。外見の欠点を隠したり、長所を強調したりするのに役立ちます。
口や足に銀色のルーブルをくわえたカエルは幸運を呼び込むお守りです! ただし、お金が「眠らない」ように、お金のお守りを寝室に置くことはお勧めできません。
タリスマンの発動
どのアーティファクトも、起動後、つまり魔法の対象をチャージした直後に魔法になります。 カエルは水の中で充電する必要があります。これを行うには、置物を水の中に1日置くだけです。 彼女の体を拭くべきではありませんが、心から話し合う必要があります。ただ、あなたが彼女に何を望んでいるのかを彼女に伝えてください。
そして彼女をお風呂に入れるのを忘れないでください!
|
これに、神経、筋肉、心臓、平滑筋、その他多くの器官や組織の機能の基本パターンを確立することができたのは、カエルでの実験だったことを付け加えることもできます。 初心者の生物学者にとって最も一般的な課題は、おそらくカエルの心臓の機能の研究でしょう。 実験プロトコルと手順は、世界中のあらゆる言語で書かれた何千ものマニュアルやワークショップに記載されています。 ただし、適切なスキルが不足しているため、実験者は最初はマニュアルに記載されているものを正確に認識しません。 私たちは、カエルの心臓の構造と機能を説明するいくつかの実際の写真とビデオクリップを閲覧して研究できるように提供しています。 すべての記録は、カエル Rana Temporaria での 1 回の実験中に行われました。 心臓の外部構造。 米。 1 カエルの心臓は胸骨の下の胸腹腔にあります。 背側、腹側、および側面(右)から見た一般的な構造を図に示します。 1:1 - 大動脈弓。 2 - 右心房。 3 - 動脈幹の下の穴に毛を挿入します。 4 - 大動脈球部(動脈円錐)。 5 - 左心房。 6 - 動脈幹。 7 - 冠状溝。 8 - 心室。 9 - 前部(頭蓋)大静脈。 10 - 肺静脈(側面図では右の静脈のみ)。 11 - 静脈洞。 12 - 後部(尾側)大静脈。 13 - 外頸静脈。 14 - 無名静脈。 15 - 鎖骨下静脈)。 両生類の心臓は、2 つの心房と 1 つの心室の 3 つの部屋で構成されていると伝統的に考えられています。 厳密に言えば、これは完全に真実ではありません。 心臓にはさらに 2 つのセクションがあり、魚類、両生類、一部の爬虫類などの下等脊椎動物では別々の部屋として明確に分離されています。 これらは、静脈洞(静脈洞)と円錐動脈(大動脈球)です。 静脈洞は、大静脈の後方 (尾側) と左右の 2 つの前方 (頭側) の融合によって形成される薄壁の部屋です。 洞は心臓の背側に位置しており、心臓の頂点を頭の方へそっと前方に引くと見ることができます。 筋肉の円錐動脈は、心室と短動脈幹(血管系の一部です)との間の腹側に位置し、そこから左右の大動脈弓が生じます。 動脈幹は心房の腹側表面に付着しておらず、その下を細い毛が通過できます(図1の剛毛「3」)。 なぜ静脈洞と円錐動脈は血管系の一部ではなく心臓の一部とみなされているのでしょうか? これは、心臓と血管の胎児の発達の特徴に基づいて行われました。 胚発生中に、静脈洞、心房、心室、動脈円錐を含む心臓はある原始から生じ、血管は別の原始から生じます。 温血動物では、発達の特定の段階では、静脈洞と動脈円錐筋もはっきりと見えます。 次に、静脈洞は右心房の壁にある洞結節(ペースメーカーゾーン、ペースメーカー)に変化し、円錐動脈は左心室と大動脈の境界にある筋肉のない輪に変化します。 したがって、温血動物の洞結節は、下等脊椎動物の静脈洞の相同物です。 ペースメーカー(英語では「pacemaker」、pacemaker)は、心臓の収縮のリズムを決定します。 自動性を備えた特殊な筋線維があります。 ペースメーカー心筋細胞では、一定の周期性を持って自発的に興奮波が発生し、心房、心室、動脈円錐体の心筋に順次伝播します。 心臓のさまざまな部分の境界(カエルでは静脈洞と心房、心房と心室、心室と動脈円錐体の間)では、励起波はより低速で伝わり、伝達には遅延が生じます。興奮の伝導が起こり、収縮波にも同じ遅延が伴います。 カエルの心臓の静脈洞は、筋肉質の洞房輪に囲まれた広い卵円孔を通って右心房と連絡しています。 洞房輪の収縮により、右心房から静脈洞への血液の逆流が部分的に防止されます。 ここには他のバルブ構造はありません。 心房は冠状溝によって心室から分離されています。 心臓は、前部 (心房、出入りする血管) と後部 (心室) に分かれています。 カエルの心臓機能の現場研究
皮弁を剥がした後、ハサミやピンセットは水ですすぎ、乾燥させてください。 米。 3 腹部の表面にあるさまざまな筋肉の中でも、腹直筋はよく目立ちます(図3)。 腹部の静脈が体軸に沿ってはっきりと見えます。 皮膚を除去した部分にカーソルを置くと、肩甲帯の骨の図(投影)が表示されます。 腹部静脈の側にある小さなピンセットを使用して、胸骨の軟骨部分 (os sternum) の端をつかみ、上方に持ち上げます。 鋭利なハサミの先端を使用して、ピンセットの隣に腹腔の筋肉壁を開きます。 注意: 胸骨の尾端近くの腹部静脈は (観察者に対して) 垂直に下向きに走っているため、それを切らないように注意する必要があります。 胸骨の端を下げずに、胸骨に沿って左右のカラス骨(os coracoideum)まで切開を続けます。 次に、大きなハサミでカラスの骨、鎖骨を切断し、胸前骨前部 (胸骨上) まで切断を続け、この筋骨格弁全体を除去します。 この後、カエルの前脚と下顎の端を穿刺して手術野を広げます。
米。 5 ビデオ クリップをオンにすると、心房と心室の連続的な収縮を観察できます ( クリップ 1)。 心房収縮期と心室収縮期の間の明確な時間遅延に注目してください。 この遅延は、心臓に部屋があるすべての動物 (脊椎動物や軟体動物) の心臓に典型的なものです。 房室遅延は心臓の効率を高めます。 この期間中、心房からの血液のほとんどが心室に移動する時間があります。 その場では、大動脈幹が心臓の基部を背側に引っ張ります。 したがって、心房の背側部分近くの心膜血管を検査するには、セルフィンカを使用して心室を尾方向にわずかに引っ張る必要があります。 サーフフックは弾性ワイヤーで作られた軽量のクランプです。 その助けを借りて、心室の頂点が注意深く掴まれます。 サーフボードの長手軸は心臓の長手軸と一致する必要があります。 まず、結紮(糸)をサーフィンのリングに結び、実験者はそれを使って心臓をキモグラフやその他のセンサーに接続します。 心臓を頭尾方向(図 6 の左側)にわずかに伸ばすと、腹側大動脈動脈系の要素をよりよく検査し、心臓の静脈部分の一部の領域を確認できるようになります(図 6)。 )。 両心房、大動脈球部、大動脈幹、大動脈弓がはっきりと見えるようになり、さらにその下に、前大静脈の基部が深く見えるようになります。 静脈床の構造のいくつかの特徴は、次の高倍率の図で見ることができます (図 7)。 図の左上の四分円では、左心房の一部が投影されていますが、ビデオカメラの焦点には入っておらず、その下には主要な静脈が見えており、左前(頭側)大静脈を形成しています。
これらの写真に付随するビデオクリップ ( クリップ 2、クリップ 3)大静脈、心房、心室、および円錐動脈の収縮の順序に注意を払う必要があります。
ビデオクリップ ( クリップ 4)小帯が原位置での心室の動きを大幅に制限していることは明らかです。 カエルの心拍をその場で記録するどの方法でも、心臓の小帯を切断する必要があります。 これは実験を成功させるために必要な条件です。 小帯と心房と大静脈の間の心膜の残骸を除去すると、静脈洞領域がはっきりと見えるようになります (図 9)。 右側では、心室が動物の体の軸に対して左(図では下向き)に移動した後の様子がはっきりと見えます(図10)。 静脈は血液で満たされており、その境界は容易に識別されます。 後大静脈は非常に短いため、実験を行う際にはこの点に注意する必要があります。 心臓を隔離するときの結紮や切断は、静脈の肝端に近づけて行う必要があります。そうしないと、静脈洞が損傷し、隔離された心臓の収縮が停止する可能性があります。
静脈洞と右心房の境界にある光の輪に注目してください。 この筋肉の輪 (洞房輪、図 9) は 2 つの部屋 (静脈洞と右心房) の間に位置し、活発に収縮し、部分的に弁の機能を果たします。 心臓の右側にある静脈洞のさまざまなセクションの収縮段階は、次の方法で分析できます。 クリップ 5)。 通常の状態では、興奮の伝播順序は、大静脈、静脈洞および心臓の近隣領域の状態によって異なります。 心臓の機能状態が変化すると、心臓の収縮のリズムを決定する一次電気活動のゾーンが移動する可能性があります。 これは通常時と外部条件が悪化した場合の両方で発生します。 生体外条件での長時間の経験により、カエルの心臓を 20 ℃以上に温めると不整脈が発生します。 まず、心房から心室への興奮の伝導が遮断され、その後、大静脈と静脈洞の収縮のみが起こります。 通常、このゾーンの一連の収縮を分析できます。 クリップ 6。 次のクリップ ( クリップ 7) は、15 分後の静脈洞と心房の機能の変化を示しています。 完全な洞房ブロックが発生しており、興奮波は心房に伝わらず、ブロックの場所は洞房輪です。 文学 ノズドラチェフ A.D.、ポリアコフ E.L. カエルの解剖学。 (実験動物)。 M.、高等学校」、1994年。P. 320。 |
 両生類は 4 本足の脊椎動物の綱に属し、この綱にはカエル、サンショウウオ、イモリなどを含む合計約 6,700 種の動物が含まれます。 このクラスは小規模とみなされます。 ロシアでは28種、マダガスカルでは247種が見つかっています。
両生類は 4 本足の脊椎動物の綱に属し、この綱にはカエル、サンショウウオ、イモリなどを含む合計約 6,700 種の動物が含まれます。 このクラスは小規模とみなされます。 ロシアでは28種、マダガスカルでは247種が見つかっています。
両生類は陸生の原始脊椎動物に属し、ほとんどの種は水生環境で繁殖および発達し、成熟した個体は陸上で生活を始めるため、水生脊椎動物と陸生脊椎動物の中間的な位置を占めます。
両生類では 肺があります、彼らが呼吸する、血液循環は 2 つの円で構成され、心臓は 3 つの部屋から構成されます。 両生類の血液は静脈と動脈に分けられます。 両生類の運動は 5 本の指の手足の助けを借りて行われ、その関節は球形です。 背骨と頭蓋骨は可動式に関節接続されています。 口蓋方形軟骨は自柱と癒合し、下顎骨は耳小骨になります。 両生類の聴覚は魚類よりも発達しており、内耳に加えて中耳もあります。 目はさまざまな距離でもよく見えるように適応しています。
両生類は陸上での生活に完全には適応していません。これはすべての器官に見られます。 両生類の温度は、環境の湿度と温度によって異なります。 彼らの陸上でのナビゲーションと移動の能力は限られています。
血液循環と循環系
 両生類 三室の心臓を持っている、心室と 2 つの心房で構成されます。 尾状動物や脚のない動物では、右心房と左心房が完全には分離されていません。 無尾類は心房間に完全な中隔を持っていますが、両生類には心室と両心房を繋ぐ共通の開口部が 1 つあります。 さらに、両生類の心臓には静脈洞があり、静脈血を受け取り、右心房と連絡しています。 円錐動脈は心臓に隣接しており、血液は心室から流れ込みます。
両生類 三室の心臓を持っている、心室と 2 つの心房で構成されます。 尾状動物や脚のない動物では、右心房と左心房が完全には分離されていません。 無尾類は心房間に完全な中隔を持っていますが、両生類には心室と両心房を繋ぐ共通の開口部が 1 つあります。 さらに、両生類の心臓には静脈洞があり、静脈血を受け取り、右心房と連絡しています。 円錐動脈は心臓に隣接しており、血液は心室から流れ込みます。
動脈円錐筋には、 スパイラルバルブ、血液を3対の血管に分配します。 心臓指数は、体重に対する心臓の質量の割合であり、動物がどれだけ活動的であるかによって決まります。 たとえば、草や緑のカエルはほとんど動かず、心臓指数は 0.5 パーセント未満です。 そして、活動的な陸生ヒキガエルの割合はほぼ 1 パーセントです。
両生類の幼生では、血液循環は 1 つの円を持ち、その血液供給システムは魚と似ています。心臓に 1 つの心房と心室があり、4 対の鰓動脈に分岐する円錐動脈があります。 最初の 3 つの動脈は外鰓と内鰓で毛細血管に分かれ、鰓毛細管は鰓動脈で合流します。 鰓枝の最初の弓を実行する動脈は、頭に血液を供給する頸動脈に分かれます。
 2回目と3回目のマージ 遠心性鰓動脈大動脈の右根と左根があり、それらの接続は背側大動脈で起こります。 鰓動脈の最後のペアは毛細血管に分かれません。これは、内鰓と外鰓につながる 4 番目の弓で、背側大動脈の根元に流れ込むためです。 肺の発達と形成は、循環の変化を伴って起こります。
2回目と3回目のマージ 遠心性鰓動脈大動脈の右根と左根があり、それらの接続は背側大動脈で起こります。 鰓動脈の最後のペアは毛細血管に分かれません。これは、内鰓と外鰓につながる 4 番目の弓で、背側大動脈の根元に流れ込むためです。 肺の発達と形成は、循環の変化を伴って起こります。
心房は縦中隔によって左右に分けられ、心臓は 3 室になっています。 毛細血管のネットワークは減少して頸動脈に変わり、背側大動脈の根元は2番目のペアから始まり、尾状では3番目のペアが保存され、4番目のペアは皮膚肺動脈に変わります。 末梢循環系も変化し、陸上系と水系の中間的な性質を獲得します。 最大の再構成は尾のない両生類で起こります。
大人の両生類には 3 つの部屋からなる心臓があります。 1つの心室と心房 2個分の量で。 壁の薄い静脈洞が右側の心房に隣接し、円錐動脈が心室から伸びています。 心臓には 5 つのセクションがあると結論付けることができます。 両方の心房が心室に通じる共通の開口部があります。 前室弁もそこに位置しており、心室が収縮するときに血液が心房に逆流するのを防ぎます。
多数の部屋が形成され、心室壁の筋肉の成長により互いに連通しています。これにより、血液が混合することができません。 右心室からは動脈円錐が伸びており、その内側には螺旋状の錐体が存在します。 3 対の動脈弓がこの円錐から離れ始めます; 最初は、血管は共通の膜を持っています。
左右の肺皮動脈まずコーンから遠ざかります。 その後、大動脈の根が現れ始めます。 2 つの鰓弓は 2 つの動脈、つまり鎖骨下動脈と後頭椎骨動脈を隔てており、前肢と体幹の筋肉に血液を供給し、脊柱の下で背側大動脈に合流します。 背側大動脈は強力な腸間膜動脈を分離しています (この動脈は消化管に血液を供給します)。 他の枝に関しては、血液は背側大動脈を通って後肢や他の臓器に流れます。
頸動脈
頸動脈は円錐動脈から最後に出発し、 内部と外部に分かれます動脈。 後肢や後部からの静脈血は坐骨静脈と大腿静脈に集まり、腎門脈に合流して腎臓で毛細血管に分かれ、腎門脈系が形成されます。 静脈は左右の大腿静脈から出て腹部奇静脈に合流し、腹壁に沿って肝臓に向かい、毛細血管に分解されます。
肝臓の門脈は胃や腸のあらゆる部分の静脈から血液を集め、肝臓で毛細血管に分割します。 腎毛細血管は静脈に合流し、遠心性で後奇大静脈に流れ込み、生殖腺から伸びる静脈もそこに流れます。 後大静脈は肝臓を通過しますが、それに含まれる血液は肝臓に入らず、肝臓からの細い静脈が肝臓に流れ込み、さらに副鼻腔静脈に流れ込みます。 すべての尾のある両生類と一部の無尾類は後基脈を保持しており、その流れは中空の前静脈に流れ込みます。
これは皮膚で酸化されて大きな皮静脈に集まり、皮静脈は静脈血を運び、上腕静脈から直接鎖骨下静脈に入ります。 鎖骨下静脈は内頸静脈および外頸静脈と合流して左前中空静脈となり、静脈洞に流れ込みます。 そこからの血液は右側の心房に流れ始めます。 肺静脈は肺からの動脈血を集め、静脈は左側の心房に流れ込みます。
動脈血と心房
 肺呼吸の場合、混合血液が右側の心房に集まり始めます。混合血液は静脈血と動脈血で構成され、静脈血は大静脈を通ってあらゆる部分から来て、動脈血は皮膚の静脈を通って来ます。 動脈血 アトリウムを埋め尽くす左側では、血液は肺から来ています。 心房の収縮が同時に起こると、血液が心室に入り、胃の壁によって血液の混合が妨げられます。右心室では静脈血が優勢となり、左心室では動脈血が優勢になります。
肺呼吸の場合、混合血液が右側の心房に集まり始めます。混合血液は静脈血と動脈血で構成され、静脈血は大静脈を通ってあらゆる部分から来て、動脈血は皮膚の静脈を通って来ます。 動脈血 アトリウムを埋め尽くす左側では、血液は肺から来ています。 心房の収縮が同時に起こると、血液が心室に入り、胃の壁によって血液の混合が妨げられます。右心室では静脈血が優勢となり、左心室では動脈血が優勢になります。
動脈円錐は右側の心室から伸びているため、心室が円錐内に収縮すると、静脈血が最初に流入し、皮膚肺動脈を満たす。 心室が錐体動脈内で収縮し続けると、圧力が上昇し始め、らせん弁が動き始め、 大動脈弓の開口部を開きます、混合血液が心室の中心から彼らに流れ込みます。 心室が完全に収縮すると、左半分からの動脈血が円錐形に入ります。
アーチ型大動脈や肺皮動脈にはすでに血液が流れているため、そこに入ることができません。強い圧力で螺旋弁が動き、頸動脈の口が開き、動脈血がそこに流れ、そこに向けられます。頭。 たとえば水中で越冬している間など、肺呼吸が長期間停止されると、より多くの静脈血が頭部に目覚めます。
代謝機能が全体的に低下し、動物は昏迷に陥るため、脳に入る酸素の量は減少します。 尾状グループに属する両生類では、両心房の間に穴が残っていることが多く、円錐動脈のらせん状の弁が十分に発達していません。 したがって、動脈弓に入る血液は、尾のない両生類よりも混合されています。
両生類であるにもかかわらず、 血液循環は2つの循環を循環します、心室が1つしかないという事実により、それらが完全に分離することはできません。 このようなシステムの構造は呼吸器官に直接関係しており、呼吸器官は二重構造を持ち、両生類のライフスタイルに対応しています。 これにより、陸上でも水中でも長い時間を過ごすことができるようになります。
赤い骨髄
両生類では長骨の赤い骨髄が現れ始めます。 総血液量は両生類の総重量の最大 7 パーセントで、ヘモグロビンは 2 ~ 10 パーセント、つまり質量 1 キログラムあたり最大 5 グラムの範囲で変化し、血液中の酸素容量は 2.5 ~ 13 の範囲で変化します。パーセントであるため、これらの数値は魚に比べて高くなります。
両生類は大きな赤血球を持っていますしかし、それらはほとんどなく、血液1立方ミリメートルあたり20万から73万です。 幼虫の血球数は成虫の血球数よりも低いです。 魚などの両生類では、血糖値は時期によって変化します。 最も高い値は、尾のある魚類と両生類で10から60パーセント、尾のない両生類では40から80パーセントで示されています。
夏が終わると、筋肉や肝臓に炭水化物が蓄積するため、冬に備えて血液中の炭水化物が大幅に増加します。また、春には繁殖期が始まり、炭水化物が血液中に流入します。 両生類は、不完全ではありますが、炭水化物代謝のホルモン調節機構を備えています。
両生類の 3 目
両生類 次のグループに分けられます。

両生類の動脈には次の種類があります。
- 頸動脈は頭に動脈血を供給します。
- 皮膚肺動脈は静脈血を皮膚と肺に運びます。
- 大動脈弓は、混合された血液を残りの臓器に運びます。
両生類は捕食者であり、唾液腺がよく発達しており、その分泌物は潤いを与えます。

両生類はデボン紀中期または下期、つまりデボン紀に発生しました。 約3億年前。 魚は彼らの祖先であり、肺があり、一対のひれがあり、そこから五本指の手足が発達した可能性が非常に高いです。 古代の葉鰭の魚はこれらの要件を満たしています。 彼らには肺があり、ヒレの骨格には、5本指の陸肢の骨格の一部に似た要素がはっきりと見えます。 また、両生類が古代の葉鰭の魚の子孫であるという事実は、古生代の両生類の頭蓋骨と類似した頭蓋骨の外皮骨の強い類似性によって示されています。
下部肋骨と上部肋骨は、葉鰭や両生類にも存在していました。 しかし、肺を持つ肺魚は両生類とは大きく異なりました。 したがって、両生類の祖先の間で陸上に行く能力を提供する運動と呼吸の特徴は、両生類が絶滅したときでも現れました。 ただの水生脊椎動物だった.
これらの適応が出現した理由は、明らかに、いくつかの種の葉鰭魚が生息する淡水貯水池の独特の体制でした。 これは、定期的な乾燥または酸素不足である可能性があります。 祖先が貯水池と決別し、陸上に定着することを決定づけた最も主要な生物学的要因は、彼らが新しい生息地で見つけた新しい食物でした。
両生類の呼吸器官
両生類は 以下の呼吸器官:

両生類では、肺は一対の袋の形で現れ、内部は空洞です。 壁の厚さは非常に薄く、内部にはわずかに発達した細胞構造があります。 しかし、両生類の肺は小さいです。 たとえば、カエルでは、皮膚に対する肺の表面の比率は、哺乳類と比較して 2 ~ 3 の比率で測定されますが、この比率は肺の方が 50 倍、場合によっては 100 倍も大きくなります。
両生類の呼吸器系の変化に伴い、 呼吸メカニズムの変化。 両生類は依然としてかなり原始的な圧力呼吸を持っています。 鼻孔を開いて口の底を下げることにより、口腔内に空気が引き込まれます。 次に、鼻孔が弁で閉じられ、空気が肺に入るために口の底が上がります。
両生類の神経系はどのように機能するのでしょうか?
両生類の脳は魚よりも重い。 脳の重量と質量の比率をパーセンテージで計算すると、軟骨を持つ現代の魚類ではこの数字は 0.06 ~ 0.44%、硬骨魚類では 0.02 ~ 0.94%、尾のある両生類では 0.29 ~ 0.36%、尾のない両生類では 0.50 ~ 0.73%。
両生類の前脳は魚類よりも発達しており、2 つの半球に完全に分かれています。 発達は、より多くの神経細胞の内容にも表れます。
脳は 5 つのセクションで構成されています。

両生類のライフスタイル
両生類が営むライフスタイルは、両生類の生理機能と構造に直接関係しています。 呼吸器官は構造が不完全です。これは主に肺に当てはまりますが、これは他の器官系に痕跡を残すためです。 肌からは常に水分が蒸発し続けますそのため、両生類は環境中の水分の存在に依存することになります。 両生類は恒温動物ではないため、生息環境の温度も非常に重要です。
このクラスの代表者は異なるライフスタイルを持っているため、構造に違いがあります。 両生類の多様性と豊富さは、湿度が高く、ほぼ常に気温が高い熱帯地方で特に高くなります。
極に近づくほど、両生類の種は少なくなります。 地球上の乾燥した寒い地域には両生類はほとんどいません。 卵は水中でのみ発育することが多いため、たとえ一時的なものであっても、水域のないところには両生類は存在しません。 塩水には両生類は存在せず、皮膚は浸透圧と高張環境を維持していません。
卵は塩水域では発育しません。 両生類は次のグループに分類されます生息地の性質により:

陸生動物は、繁殖期以外は水域から遠く離れた場所に移動することがあります。 しかし、水生動物は逆に、一生を水の中、または水のすぐ近くで過ごします。 尾のあるカエルの中では水生のカエルが優勢であり、尾のないカエルのいくつかの種もそれらに属する可能性があり、たとえばロシアでは、これらは池または湖のカエルです。
樹上両生類カイアシ類やアマガエルなどの陸生動物に広く分布しています。 陸生両生類の中には、穴を掘って生活するものもいます。たとえば、尾のないものや、ほとんどすべてが脚のないものもあります。 一般に、陸上に住む人々は肺がより発達しており、皮膚は呼吸過程にあまり関与しません。 このため、彼らが住んでいる環境の湿度にあまり依存しません。
両生類は、その数に応じて年ごとに変動する有用な活動に従事します。 特定の段階、特定の時間、特定の気象条件では異なります。 両生類は鳥以上に、味や匂いの悪い昆虫や保護色の昆虫を駆除します。 ほぼすべての食虫鳥が眠っているときに、両生類は狩りをします。
科学者たちは、両生類が菜園や果樹園の害虫駆除者として大きな利益をもたらすという事実に長い間注目してきました。 オランダ、ハンガリー、イギリスの庭師たちは特別に各国からヒキガエルを持ち込み、温室や庭に放した。 1930 年代半ば、約 150 種のアギヒキガエルがアンティル諸島とハワイから輸出されました。 彼らは繁殖を始め、100万匹以上のヒキガエルがサトウキビ農園に放たれ、その結果はすべての予想を上回りました。
 両生類の目は目詰まりや乾燥から守っています。 可動式の下まぶたおよび上まぶた、瞬膜と同様に。 角膜は凸面になり、水晶体はレンズ状になりました。 基本的に、両生類は動く物体を見ます。
両生類の目は目詰まりや乾燥から守っています。 可動式の下まぶたおよび上まぶた、瞬膜と同様に。 角膜は凸面になり、水晶体はレンズ状になりました。 基本的に、両生類は動く物体を見ます。
聴覚器としては、耳小骨と中耳が登場しました。 空気環境は水よりも密度が高いため、このように見えるのは、音の振動をよりよく知覚する必要があるためです。
循環系
準備の上部、肺と肝臓の間、薄膜で形成された心膜腔 - 心膜嚢内に心臓があります。 時々、開いた後もゆっくりと収縮し続けることがあります。 心臓の上部にある心膜嚢の薄い無色の膜をピンセットで剥がし、心臓を傷つけないように慎重にハサミで切ります。 心臓がバッグから滑り落ちます。 心臓を開かないで、後ろ向きの頂点をピンセットで持ち上げます。 2 つの大きな前後大静脈の融合によって形成された、明確な境界のない暗い薄壁の静脈洞 (洞静脈洞) が見えます。 心臓の上部には、互いに完全に分離された大きな右心房があります。 静脈洞がその中に開口しています。 外側では、心房間の境界は非常に弱く表現されます。 心臓の下部のピンクがかった円錐形の最も筋肉の部分が見えます。これが心室です。 両方の心房は共通の開口部を介して心房と通信します。 この穴の特別な弁により、血液は心房から心室への一方向にのみ流れることができます。
動脈血はまばらな陰影で示され、混合血液は濃い陰影で示され、静脈血は黒色で示されます。
5 - 総動脈幹、6 - 肺皮動脈、7 - 肺動脈、
8 - 大きな皮動脈、9 - 右大動脈弓、10 - 左大動脈弓、
11 - 後頭椎骨動脈、12 - 鎖骨下動脈、13 - 背側大動脈、
14 - 腸間膜動脈、15 - 尿生殖器動脈、16 - 総腸骨動脈、17 - 総頚動脈、18 - 内頚動脈、19 - 外頚動脈、20 - 頚動脈、21 - 肺、22 - 肝臓、23 - 胃、24 - 腸、
25 - 精巣、26 - 腎臓
円錐動脈は心室の右側から発生します。 したがって、両生類の心臓は 3 室 (2 つの心房と 1 つの心室) ですが、静脈洞、2 つの心房、心室、円錐動脈という 5 つのセクションで構成されています。 円錐動脈は 3 対の動脈弓を生じます。 各アーチは独立した開口部を持って動脈円錐から離れています。 左側と右側の 3 つのアーチはすべて、最初は共通の膜で囲まれた共通の動脈幹と一体になっているため、動脈円錐が 2 つの大きな幹にのみ分割されているような印象が生じます。 しかし、この幹を針で持ち上げると、別々ではあるが隣接した血管で構成されていることがはっきりとわかります。 これらの血管(動脈円錐筋からの起源の順)は次のとおりです。
1. 動脈円錐の背側部分から最初に離れるのは、一対の(右と左)皮膚肺動脈です。これは、魚のIV対の鰓動脈弓の相同体です。 すぐに、各肺皮動脈は、肺の端に沿って肺尖まで走る肺動脈と、体の背側表面の皮膚で分岐する大きな皮動脈に分かれます。
2. 皮膚肺動脈のすぐ後ろですが、円錐動脈の腹部から、対の大動脈弓が生じます。 それらは、鰓動脈弓の 2 番目のペアと相同です。 各大動脈弓は上方(体の背側表面に向かって)および側方に湾曲し、後頭椎弓と鎖骨下弓を分けています。 動脈の前肢に血液を供給します。 その後、大動脈弓は脊柱の下 (胃の後ろの高さ) で互いに結合し、奇突起背側大動脈を形成します。 強力な腸間膜動脈は背側大動脈から出発します。 それは腸間膜のひだに沿って通過し、血液を胃、腸、肝臓、脾臓に運びます。 逆向きに走る背側大動脈(そこから出発した腸間膜動脈よりも細い幹のように見える)は、腎臓と生殖器に細い動脈を数本出しています。 背側大動脈は腎臓の後端のレベルで 2 つの総腸骨動脈に分かれ、その枝は体の後部と後肢に血液を供給します。
3. 頭部に血液を供給する頸動脈弓 (頸動脈) は、大動脈弓に沿って、また総頸動脈の形で動脈円錐の腹部からも伸びています。 総動脈幹を出たほぼ直後に、各頸動脈弓は外頸動脈と内頸動脈に分かれます。 内動脈の分岐部位、つまり内動脈の基部には頸動脈、つまり頸動脈の「腺」があり、頸動脈内の血圧を調節しているようです。

.
静脈血は黒く表示されますが、
動脈 - ハッチング付き、混合 - ドット付き:
1 - 静脈洞、2 - 右心房、3 - 左心房、4 - 心室、
5 - 外頸静脈、6 - 内頸静脈、7 - 大皮静脈、
8 - 上腕静脈、9 - 鎖骨下静脈、10 - 右前大静脈、
11 - 左前大静脈、12 - 大腿静脈、13 - 坐骨静脈、
14 - 総腸骨静脈、または腎臓の門脈、15 - 腹部静脈、
16 - 肝臓の門脈、17 - 遠心性腎静脈、18 - 後大静脈、
19 - 肝静脈、20 - 肺静脈、21 - 肺、22 - 肝臓、23 - 胃、
24 - 腸、25 - 精巣、26 - 腎臓
頭部からの静脈血は外頸静脈と内頸静脈を通過します。 皮膚で酸化された動脈血は強力な太い皮静脈を流れ、そこに前肢から静脈血を運ぶ上腕静脈が流れ込みます。 皮膚静脈と上腕静脈は鎖骨下静脈に合流します。 ほぼ即座に、両側の鎖骨下静脈が外頸静脈と内頸静脈と合流し、左右の前大静脈を形成します。 動脈血と混合した静脈血を運ぶ(大皮静脈を通って入る)両方の前大静脈は、静脈洞に流れ込みます。
後肢と骨盤領域から、静脈血がいくつかの静脈を通って流れます。 それらの中で最大のものは大腿静脈と坐骨静脈で、両側で合流して対の総腸骨静脈、または腎門脈を形成し、腎臓に入り、そこで毛細血管のネットワーク(腎門脈系)に分かれます。
幹は左右の大腿静脈から出ており、互いに合流して腹部静脈を形成します。 それは体の腹壁に沿って通過し、膀胱と筋肉から血液を集め、胸骨の後端近くで腹腔に突入し、肝臓に進み、そこで毛細血管に分裂します。 腸、胃、食道のあらゆる部分からの静脈血は、静脈系を通って肝臓の大きな門脈に集められ、肝臓に入り、そこで毛細血管に分かれます。 したがって、両生類では、肝臓の門脈系は肝臓自体の門脈と腹部静脈の2つの静脈によって形成されています。
腎臓の毛細血管を通過する静脈血は、いくつかの遠心性腎静脈に集まり、奇静脈後大静脈に合流します。 生殖腺から血液を運ぶ静脈がそこに流れ込みます。 後大静脈はすぐに肝臓の中央部に入り、肝臓を貫通します(そこからの血液は肝臓に入りません)。 後大静脈は肝臓からの出口で 2 本の短い肝静脈を受け取り、静脈洞に流れ込みます。
肺からの動脈血は肺静脈を通って流れ、肺静脈は合流して左心房に流れ込みます。 それらの合流点は左前大静脈で覆われています。
両生類では、小さい血液循環(心室 - 肺 - 左心房)と大きい血液循環(心室 - 全身 - 静脈洞 - 右心房)の 2 つの血液循環が明確に表現されます。 ただし、心室が 1 つであるため、これらの円はまだ完全に分離されておらず、血液の一部が心室内で混ざります。 両生類の活動状態では、肺と皮膚の両方で血液の酸素飽和が起こります。 したがって、右心房には、静脈血(全身の静脈から静脈洞に集められる)と動脈血(大きな皮静脈によってもたらされる)が混合されています。 左心房では、血液は動脈血です(肺から肺静脈を通って来た)。 心房も同時に収縮し、血液が心室に入ります。 筋肉の成長が強力に発達しているため、心室の空洞は、いわば多数の部屋に分割されており(図48)、血液の混合を妨げます。 したがって、心室の右部分には動脈血と混合した静脈血(右心房と同じ組成)があり、心室の左部分には動脈血(左心房と同じ)があり、真ん中部分は血が混ざっています。

:
1 - 右心房、2 - 左心房、3 - 心室、4 - 弁、
両心房から心室につながる共通の開口部を閉じる、
5 - 動脈円錐、6 - 総動脈幹、7 - 肺皮動脈、
8 - 大動脈弓、9 - 総頸動脈、10 - 頸動脈、
11 - 動脈円錐のスパイラル弁
心室から動脈幹への血液の流れはまだ十分に研究されていません。 このプロセスは次のように簡略化できます。 心室が収縮すると、より多くの静脈血がまず動脈円錐形筋に流入します(その起源が心室の右側からであるため)。 それは開いた開口部を通って直ちに皮膚肺動脈を満たし(残りの動脈弓の開口部は動脈円錐の螺旋弁によって閉じられている)、肺および皮膚に入り酸化される。 継続的な心室収縮によって肺皮動脈が満たされた後、円錐動脈内の圧力が増加します。 らせん弁が動き、大動脈弓の開口部が開きます。 混合血液は心室の中央部からそれらに突入し、大動脈弓の枝と背側大動脈の枝に沿って体全体に広がります。
心室の最大収縮中に心室の左側から円錐動脈に出る動脈血は、すでに血液で満たされているため、皮膚肺動脈および大動脈弓に入ることができません。 螺旋弁が最大に移動し、頸動脈の口が解放されます。 それらを通って、動脈血は頭部(脳および感覚器官を含む)に送られます。
尾のない両生類において、肺呼吸が長期間停止しているとき(たとえば、皮膚の表面でのみ呼吸が行われる貯水池の底で越冬しているとき)における血流の分離のメカニズムはまだ解明されていない。 尾状両生類では、円錐動脈の螺旋弁が十分に発達していないため、より多くの混合血液がすべての動脈弓に流入します。
呼吸器系
呼吸器系には、経路と肺の両方が含まれます。 喉頭裂は、いくつかの軟骨によって口腔から区切られ、小さな空洞である喉頭(喉頭)に通じています。 喉頭裂は、喉頭の特別な筋肉の収縮によって開閉できます。 披裂軟骨の凹面の内側には声帯、つまり喉頭の粘膜のひだがあります。 空気が喉頭を通過することによってこれらの靭帯が振動すると、共鳴器によって増幅された音(きしみ音)が発生します。 喉頭腔からの 2 つの小さな開口部は、対の肺に直接つながっています。

:
1 - 右心房、2 - 左心房、3 - 心室、4 - 円錐動脈、
5 - 肺、6 - 食道、7 - 胃、8 - 胃の幽門部、
9 - 十二指腸、10 - 膵臓、11; -小腸、
12 - 直腸、13 - 総排出腔領域、14 - 肝臓、15 - 胆嚢、
16 - 胆管、17 - 腸間膜、18 - 脾臓、19 - 腎臓、20 - 尿管、
21 - 膀胱、22 - 卵巣、23 - 卵管
(左卵巣と卵管は図には示されていません)
心臓の側面にある肺は、弾性壁を備えた薄壁の袋であり、外側は細胞構造をしています(肺をよりよく検査するには、細い端を喉頭スリットに挿入したガラス管を通して肺をわずかに膨らませる必要があります)。 細胞性は、肺の壁の内側にある小さな増殖物(隔壁)によって引き起こされ、これにより、その内面がわずかに増加します。 しかし、両生類の肺の内面の総面積は小さく、通常は皮膚の表面よりも若干小さい(いくつかの種ではわずかに大きい)ことさえあります。 (哺乳類では、肺の内面は皮膚の表面の 60 ~ 100 倍も大きくなります。)
実際の陸生脊椎動物(爬虫類、鳥類、哺乳類)では、空気は主に胸部(肋骨を介した胸骨と柱との接続部分)の容積を変化させることによって肺に吸い込まれます。 両生類には胸がありません(肋骨がないか、または非常に発達が不十分です)、そしてその呼吸メカニズムは非常に独特です。 生きているカエルでは、口の底がリズミカルに上がったり下がったりするのがはっきりとわかります。 鼻孔の外側の開口部は、異なるリズムで開閉します。

.
I - 口腔が拡張し、開いた鼻孔から空気が入ります。
II - 鼻孔が閉じ、喉頭スリットが開き、肺から出た空気が口腔内で大気と混合します。 III - 鼻孔が閉じている、
口腔が収縮し、混合された空気が肺に押し込まれます。
IV - 喉頭裂を閉じ、口腔の底を口蓋に押し付けます。
残った空気を開いた鼻孔から押し出します。
1 - 鼻孔の外部開口部、2 - 鼻孔の内部開口部(チョアナ)、3 - 口腔
空洞、4 - 口腔底、5 - 喉頭裂、6 - 肺、7 - 食道
口腔の底が下がると、口腔の体積が大幅に増加し、空気が鼻道(開いた外鼻孔と鼻腔)を通って口腔内に吸い込まれます。 この時点では喉頭裂は閉じています。 次に、外鼻孔が閉じます(これは鼻孔の特別な筋肉の作用によって起こります。位置を変える上顎前骨の突起も閉じるのに役立ちます)、同時に喉頭裂が開きます。 空気は肺から(内臓からの圧力と腹壁の筋肉の収縮を受けて)膨張し続ける口腔に入り、そこにある大気と混合します。
次に、口腔の底が口蓋に向けて徐々に上昇し始め、口腔から混合された空気が肺に押し込まれます。 次に、喉頭のスリットが閉じ、口腔の底が口蓋に押し付けられ、残った混合空気が開いた鼻孔から押し出されます。 その後、吸入の最初の段階が再び始まります。
不規則な呼吸運動の合間に、口腔底は鼻孔が開き、喉頭スリットが閉じた状態で小さな振幅の振動を起こします。 同時に、口腔内の空気が更新され、口腔粘膜の毛細血管内の血液が酸素で飽和されます。
消化器系
消化管は口腔から始まり総排出腔で終わります。 喉頭裂の後ろの口腔から、短くて容易に伸長可能な食道が広い開口部で始まり、心臓、肺、肝臓の上の体腔の背側に沿って通過します。 肝臓の右葉(開口部から)を左に回すと、その後ろの部分がはっきりと見えます。
食道はリング状の狭窄部によって胃から分離されて胃に流れ込みます。 胃はわずかに湾曲しており、食道よりも厚い筋肉の壁を持っています。 胃の後部の狭くなった端は、ほとんど目立たない環状の狭窄部によって、胃の前方に平行に延びる小腸の最初の部分である十二指腸から分離されています。 胃と十二指腸の間の腸間膜に、膵臓はゆるやかな黄色がかった紐の形で存在します。 鋭い境界がなく、十二指腸は体腔の右側(左側の開口部から)にいくつかのループの形で位置するわずかに小さい直径を持つ小腸に入ります。 小腸は短くて広い直腸に入り、薄くなって背側に開口部のある総排泄腔を形成します。
肝臓は大きくてコンパクトな 3 葉の臓器で、心臓のすぐ後ろにあります。 その小さな中葉の下面には、円形の緑がかった黒色の胆嚢があります。肝臓から分泌された胆汁が蓄積する貯留槽です。 肝管は肝臓の右葉と左葉の背側表面に沿って通り(注意深く検査した場合のみ見ることができます)、胆嚢管(特別な準備がなければ見ることができません)と一緒に総胆管に入ります。 密なコードの形をした胆管は、膵臓の組織を通過し、そこからいくつかの小さな短い管を受け、十二指腸の最初の部分に流れ込みます。 肝臓、胃、および腸全体は、体腔の背側表面から薄い半透明の折り畳まれたフィルム、つまり腸間膜上に吊り下げられています。
食道の機能は食べ物を胃に運ぶことです。 ここで、食べ物は筋肉の壁の圧力によって押しつぶされ、胃の腺から分泌される消化酵素に浸されます。 小腸では、食物塊に肝臓と膵臓から胆管を通って入ってくる酵素が含浸され、消化され、腸壁から吸収されます。 直腸では、水分が吸収され、便が形成され、総排出腔を通って放出されます。
小腸の腸間膜には(開いたカエルの上、通常は胃と直腸の間に)、細網内皮系の非常に重要な器官である小さな丸い脾臓があります。 脾臓では、形成された血液要素(赤血球、リンパ球)の形成が起こり、血液に入る細菌の食作用などが起こります。脾臓は、必要に応じて血液の貯蔵庫としても機能します(失血、長時間の激しい動きなど)。 )血流に放出されます。
泌尿器系
一対のコンパクトな細長い楕円形の中腎 (または幹) 腎臓は、体腔の後部の脊柱の側面に位置しています。 各腎臓の外縁に沿って、薄い灰色がかったコード、つまりウォルフ管、つまり中腎腎臓の尿管が走っています。 左右の腎臓のウォルフ管は、総排出腔の背側に独立した開口部を持って開口しています。 総排泄腔の腹壁にある不対の開口部は、大きな 2 葉の薄壁膀胱につながっています。 ピペットを使って総排出腔を通って膀胱の開口部に水を導入すると、膀胱壁の強い伸展性を簡単に確認できます。 ウォルフ管(尿管)を通って総排出腔に入った尿は、総排出腔の底に流れて膀胱に入ります。 その壁にはかなり密な毛細管ネットワークがあり、尿からの水分が確実に吸収されます。 尿はより濃縮され、膀胱壁の収縮が再び総排出腔内に排出され、総排出腔から排出されます。

:
1 - 腎臓、2 - 尿管(精管としても知られる)、3 - 総排出腔、
4 - 泌尿生殖器の開口部、5 - 膀胱、6 - 膀胱の開口部、
7 - 精巣、8 - 精細管、9 - 精嚢、
10 - 脂肪体、11 - 副腎
これらの特徴は、男女両方の排泄システムの特徴です。
各腎臓の前端、生殖腺と同じ腸間膜には、指の形をしたオレンジ色の脂肪体があり、生殖細胞の形成のための栄養素の貯蔵庫です。 各腎臓の表面に沿って、狭く、時にはかすかに目立つ黄色がかった帯、つまり副腎、内分泌腺が伸びています。
精巣は対になっており、丸く、黄色がかった色または茶色がかった色で、腎臓の前端近くの脂肪体とともに腸間膜に浮遊しています。 ピンセットで精巣を注意深く引っ張ると、精巣から伸びる細い白っぽい糸が腸間膜(腎臓の前部に流れ込む精細管)で目立ちます。 精細管は尿細管に開口しており、したがって両生類の腎臓の前部は機能的に精巣上体として機能し、雄の両生類のウォルフ管は同時に尿管と精管の両方として機能します。 繁殖期(4月、5月)には、つぼみから出た直後に、ポケット状の拡張物(精嚢)がヴォルフ管の壁にはっきりと見えます。 それらは精液の貯蔵庫として機能します。 繁殖期以外では、精嚢のサイズは減少しますが、依然として目で見ることができます。

1 - 腎臓、2 - 尿管、3 - 総排出腔、4 - 尿道口、5 - 膀胱、
6 - 膀胱の開口部、7 - 左卵巣(右卵巣は図には示されていない)、
8 - 卵管、9 - 卵管の漏斗、10 - 脂肪体(右側の脂肪体は示されていない)、11 - 副腎、12 - 生殖口(卵管の開口部)
対になった卵巣は腸間膜上に(脂肪体とともに)浮遊する薄壁の嚢であり、季節に応じて体腔の多かれ少なかれ重要な部分を満たしています。 卵巣を埋める色素のついた卵が、卵巣の壁を通してはっきりと見えます。 女性の生殖管は一対の卵管、つまり体腔の側面の短い腸間膜に吊り下げられたミュラー管です。 卵管の長さは季節によって大きく異なります。 繁殖期の春には特に複雑に伸びます。 各卵管の前端は、拡張した開口部である卵管漏斗を介して体腔 (心臓の隣) に開口しています。 卵管の下部(子宮部分と呼ばれることが多い)は幅が広くなります。 各卵管は独立した開口部で総排泄腔に通じています
1 - 総排出腔の外部開口部、2 - 総排出腔の空洞。 3 - 直腸、
4 - 膀胱、5 - 尿管、6 - 卵管、7 - 体壁
卵子が成熟すると、周囲の卵胞膜が破裂し、卵子が体腔内に押し込まれます。 ここで、それは卵管漏斗によって拾われ、産卵の開始時に急激に増加し、その壁の蠕動運動のおかげで卵管に沿って移動します。 この場合、卵管の壁にある腺の分泌物が卵子の周りに透明なゼラチン状の卵管を形成します。 卵管の下部(子宮)では、完全に形成された卵(卵子)が、産卵の準備ができた塊にまとめられます。
したがって、両生類(典型的な無羊症)では、軟骨魚と同様に、成体になっても中腎型の腎臓が機能します。 ヴォルフの雄では、この管が尿管と精管の両方の役割を果たし、精子が体腔に落ちることはなく、ミュラー管は縮小しています。 ウォルフ管のメスでは、管は尿管としてのみ機能し、ミュラー管は卵管として機能します。 成熟した卵子は体腔に落ち、漏斗を通って卵管に入ります。
末梢神経系
すべての器官系をスケッチした後、解剖したカエルの内部を取り除くと、脊柱から伸びている密な白いコード、つまり脊髄神経がはっきりと見えるようになります。 脊柱の側面には、リンパ管を通って内耳の被膜の膜迷路の空洞につながっている白い「石灰質の嚢」が見えます。 これらの嚢の機能的重要性はまだ明らかになっていません。
カエルの胎生後発達
湿った標本でカエルの幼虫、オタマジャクシの発育段階を調べます。 卵から孵化した直後のオタマジャクシには、外えらと小さな尾があります。 オタマジャクシが成長するにつれて、革のようなひだ (「鰓蓋」) が発達して外鰓を覆い、尾が成長します。 変態は徐々に進行します。四肢の基礎が現れ(後肢と同時に形成される前肢の基礎は、外えらを覆う革のようなひだで覆われているため見えません)、成長と形成が進みます。 。 尾は徐々に溶け始めます。 こうした外部の変化と同時に、消化器系、呼吸器系、循環器系といった内臓の再構築が起こります。 変態の結果、水生幼生は水の外で生きることができる小さなカエルに変わります。
トピックに関する記事